

Hepatitis B virus morphogenesis
- PMID: 17206755
- PMCID: PMC4065877
- DOI: 10.3748/wjg.v13.i1.65
Hepatitis B virus morphogenesis
Abstract
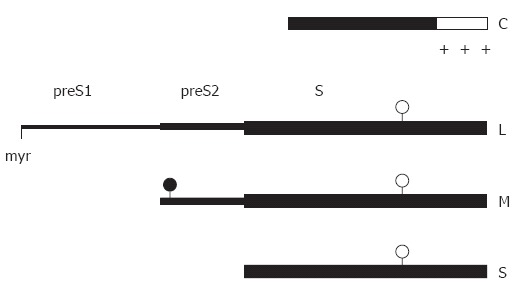
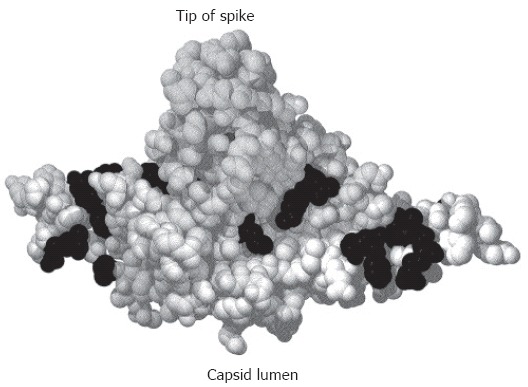
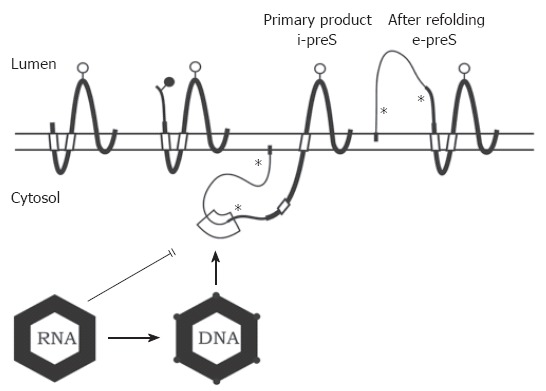
The hepatitis B virus (HBV) particle consists of an envelope containing three related surface proteins and probably lipid and an icosahedral nucleocapsid of approximately 30 nm diameter enclosing the viral DNA genome and DNA polymerase. The capsid is formed in the cytosol of the infected cell during packaging of an RNA pregenome replication complex by multiple copies of a 21-kDa C protein. The capsid gains the ability to bud during synthesis of the viral DNA genome by reverse transcription of the pregenome in the lumen of the particle. The three envelope proteins S, M, and L shape a complex transmembrane fold at the endoplasmic reticulum, and form disulfide-linked homo- and heterodimers. The transmembrane topology of a fraction of the large envelope protein L changes post-translationally, therefore, the N terminal domain of L (preS) finally appears on both sides of the membrane. During budding at an intracellular membrane, a short linear domain in the cytosolic preS region interacts with binding sites on the capsid surface. The virions are subsequently secreted into the blood. In addition, the surface proteins can bud in the absence of capsids and form subviral lipoprotein particles of 20 nm diameter which are also secreted.
Figures
References
-
- Roingeard P, Sureau C. Ultrastructural analysis of hepatitis B virus in HepG2-transfected cells with special emphasis on subviral filament morphogenesis. Hepatology. 1998;28:1128–1133. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
