

The grapevine transcription factor VvMYBPA1 regulates proanthocyanidin synthesis during fruit development
- PMID: 17208963
- PMCID: PMC1820911
- DOI: 10.1104/pp.106.093203
The grapevine transcription factor VvMYBPA1 regulates proanthocyanidin synthesis during fruit development
Abstract
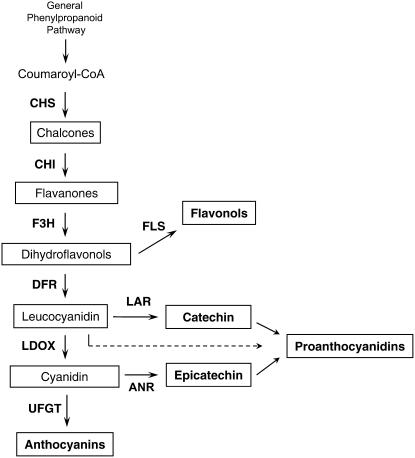
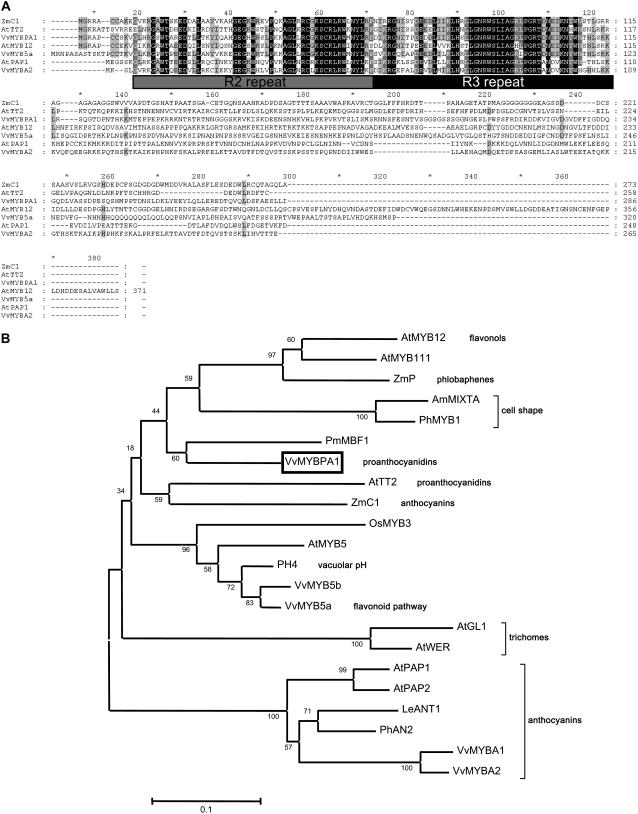
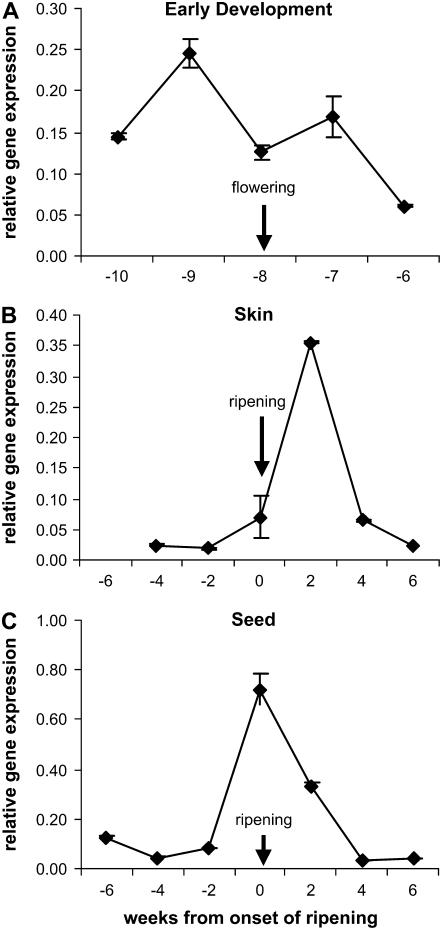
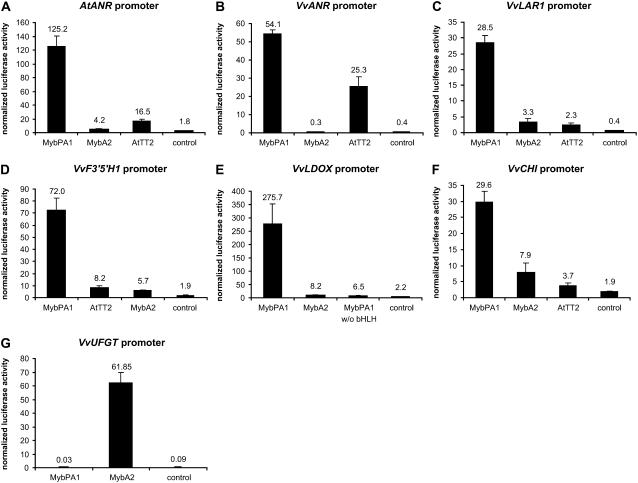
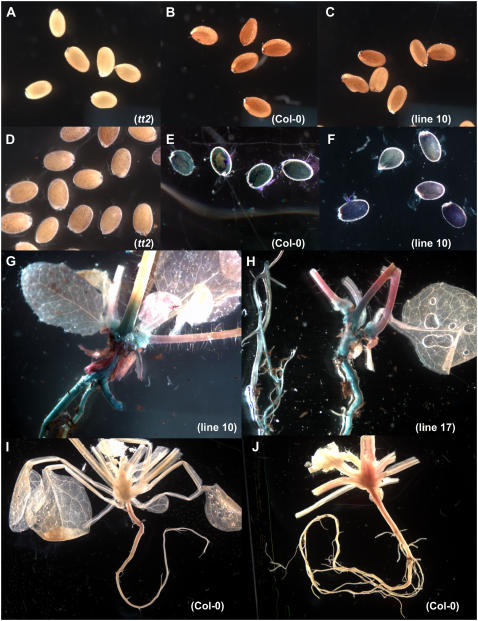
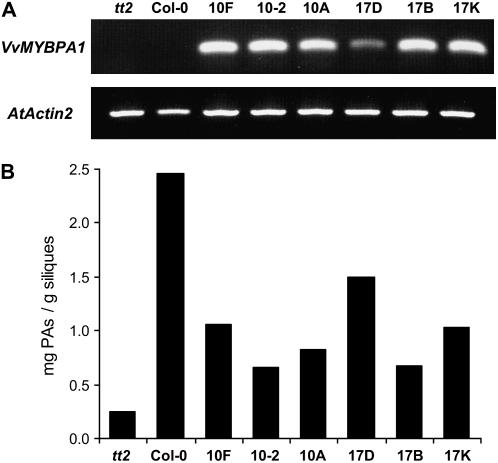
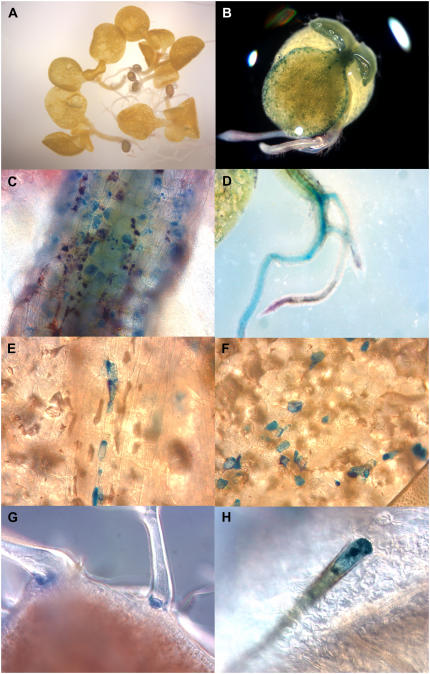
Proanthocyanidins (PAs; or condensed tannins) can protect plants against herbivores, contribute to the taste of many fruits, and act as dietary antioxidants beneficial for human health. We have previously shown that in grapevine (Vitis vinifera) PA synthesis involves both leucoanthocyanidin reductase (LAR) and anthocyanidin reductase (ANR). Here we report the characterization of a grapevine MYB transcription factor VvMYBPA1, which controls expression of PA pathway genes including both LAR and ANR. Expression of VvMYBPA1 in grape berries correlated with PA accumulation during early berry development and in seeds. In a transient assay, VvMYBPA1 activated the promoters of LAR and ANR, as well as the promoters of several of the general flavonoid pathway genes. VvMYBPA1 did not activate the promoter of VvUFGT, which encodes the anthocyanin-specific enzyme UDP-glucose:flavonoid-3-O-glucosyltransferase, suggesting VvMYBPA1 is specific to regulation of PA biosynthesis in grapes. The Arabidopsis (Arabidopsis thaliana) MYB transcription factor TRANSPARENT TESTA2 (TT2) regulates PA synthesis in the seed coat of Arabidopsis. By complementing the PA-deficient seed phenotype of the Arabidopsis tt2 mutant with VvMYBPA1, we confirmed the function of VvMYBPA1 as a transcriptional regulator of PA synthesis. In contrast to ectopic expression of TT2 in Arabidopsis, constitutive expression of VvMYBPA1 resulted in accumulation of PAs in cotyledons, vegetative meristems, leaf hairs, and roots in some of the transgenic seedlings. To our knowledge, this is the first report of a MYB factor that controls genes of the PA pathway in fruit, including both LAR and ANR, and this single MYB factor can induce ectopic PA accumulation in Arabidopsis.
Figures
References
-
- Abrahams S, Lee E, Walker AR, Tanner GJ, Larkin PJ, Ashton AR (2003) The Arabidopsis TDS4 gene encodes leucoanthocyanidin dioxygenase (LDOX) and is essential for proanthocyanidin synthesis and vacuole development. Plant J 35 624–636 - PubMed
-
- Bagchi D, Bagchi M, Stohs SJ, Das DK, Ray SD, Kuszynski CA, Joshi SS, Pruess HG (2000) Free radicals and grape seed proanthocyanidin extract: importance in human health and disease prevention. Toxicology 148 187–197 - PubMed
-
- Baudry A, Heim MA, Dubreucq B, Caboche M, Weisshaar B, Lepiniec L (2004) TT2, TT8, and TTG1 synergistically specify the expression of BANYULS and proanthocyanidin biosynthesis in Arabidopsis thaliana. Plant J 39 366–380 - PubMed
-
- Baxter IR, Young JC, Armstrong G, Foster N, Bogenschutz N, Cordova T, Peer WA, Hazen SP, Murphy AS, Harper JF (2005) A plasma membrane H+-ATPase is required for the formation of proanthocyanidins in the seed coat endothelium of Arabidopsis thaliana. Proc Natl Acad Sci USA 102 2649–2654 - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
