

Insecticidal toxins from black widow spider venom
- PMID: 17210168
- PMCID: PMC2517654
- DOI: 10.1016/j.toxicon.2006.11.021
Insecticidal toxins from black widow spider venom
Abstract
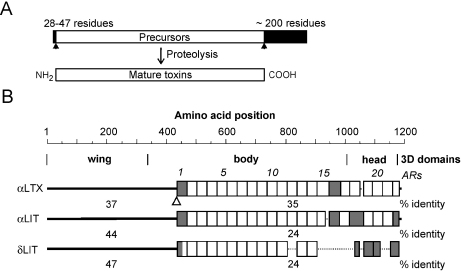
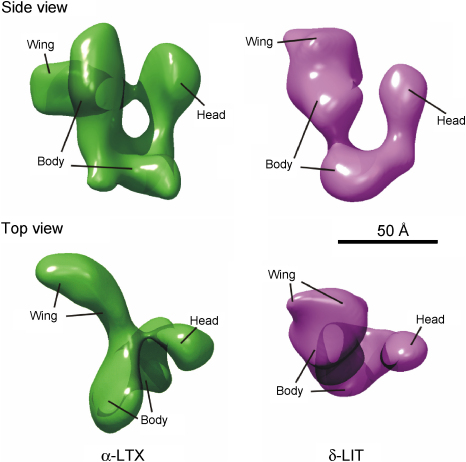
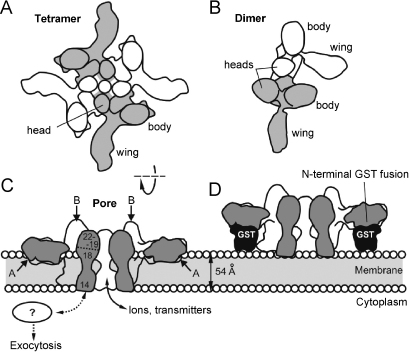
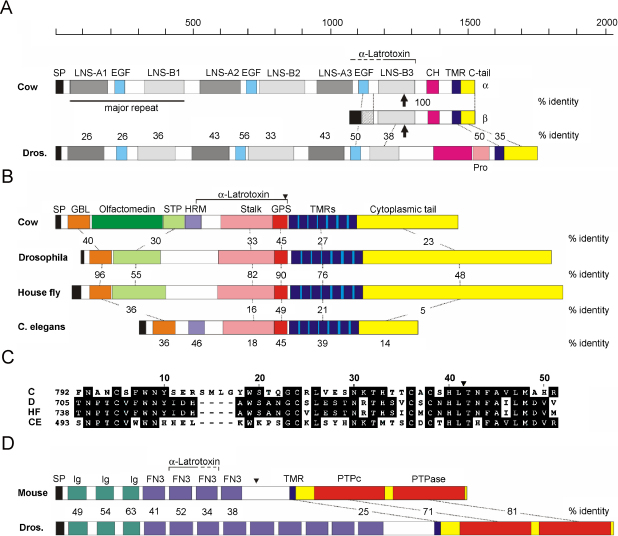
The biological effects of Latrodectus spider venom are similar in animals from different phyla, but these symptoms are caused by distinct phylum-specific neurotoxins (collectively called latrotoxins) with molecular masses ranging from 110 to 140 kDa. To date, the venom has been found to contain five insecticidal toxins, termed alpha, beta, gamma, delta and epsilon-latroinsectotoxins (LITs). There is also a vertebrate-specific neurotoxin, alpha-latrotoxin (alpha-LTX), and one toxin affecting crustaceans, alpha-latrocrustatoxin (alpha-LCT). These toxins stimulate massive release of neurotransmitters from nerve terminals and act (1) by binding to specific receptors, some of which mediate an exocytotic signal, and (2) by inserting themselves into the membrane and forming ion-permeable pores. Specific receptors for LITs have yet to be identified, but all three classes of vertebrate receptors known to bind alpha-LTX are also present in insects. All LTXs whose structures have been elucidated (alpha-LIT, delta-LIT, alpha-LTX and alpha-LCT) are highly homologous and have a similar domain architecture, which consists of a unique N-terminal sequence and a large domain composed of 13-22 ankyrin repeats. Three-dimensional (3D) structure analysis, so far done for alpha-LTX only, has revealed its dimeric nature and an ability to form symmetrical tetramers, a feature probably common to all LTXs. Only tetramers have been observed to insert into membranes and form pores. A preliminary 3D reconstruction of a delta-LIT monomer demonstrates the spatial similarity of this toxin to the monomer of alpha-LTX.
Figures
Similar articles
-
Molecular architecture of black widow spider neurotoxins.Nat Commun. 2021 Nov 29;12(1):6956. doi: 10.1038/s41467-021-26562-8. Nat Commun. 2021. PMID: 34845192 Free PMC article.
-
Expression of Brown and Southern Black Widow Spider (Araneae: Theridiidae) Latrotoxins Is Tissue- and Life Stage-Specific for α-Latroinsectotoxins and δ-Latroinsectotoxins and Is Ubiquitous for α-Latrotoxins.J Med Entomol. 2022 Jan 12;59(1):184-191. doi: 10.1093/jme/tjab168. J Med Entomol. 2022. PMID: 34632517
-
Cloning and activity of a novel α-latrotoxin from red-back spider venom.Biochem Pharmacol. 2012 Jan 1;83(1):170-83. doi: 10.1016/j.bcp.2011.09.024. Epub 2011 Oct 5. Biochem Pharmacol. 2012. PMID: 22001442
-
The multiple actions of black widow spider toxins and their selective use in neurosecretion studies.Toxicon. 2004 Apr;43(5):527-42. doi: 10.1016/j.toxicon.2004.02.008. Toxicon. 2004. PMID: 15066411 Review.
-
Mechanistic insights on spider neurotoxins.EXS. 2010;100:293-315. doi: 10.1007/978-3-7643-8338-1_8. EXS. 2010. PMID: 20358687 Review.
Cited by
-
Temporal characteristics of vesicular fusion in astrocytes: examination of synaptobrevin 2-laden vesicles at single vesicle resolution.J Physiol. 2011 Sep 1;589(17):4271-300. doi: 10.1113/jphysiol.2011.210435. Epub 2011 Jul 11. J Physiol. 2011. PMID: 21746780 Free PMC article.
-
Recent Advances in Research on Widow Spider Venoms and Toxins.Toxins (Basel). 2015 Nov 27;7(12):5055-67. doi: 10.3390/toxins7124862. Toxins (Basel). 2015. PMID: 26633495 Free PMC article. Review.
-
A tentacle for every occasion: comparing the hunting tentacles and sweeper tentacles, used for territorial competition, in the coral Galaxea fascicularis.BMC Genomics. 2020 Aug 8;21(1):548. doi: 10.1186/s12864-020-06952-w. BMC Genomics. 2020. PMID: 32770938 Free PMC article.
-
Improving the Annotation of the Venom Gland Transcriptome of Pamphobeteus verdolaga, Prospecting Novel Bioactive Peptides.Toxins (Basel). 2022 Jun 15;14(6):408. doi: 10.3390/toxins14060408. Toxins (Basel). 2022. PMID: 35737069 Free PMC article.
-
Pore-Forming Proteins from Cnidarians and Arachnids as Potential Biotechnological Tools.Toxins (Basel). 2019 Jun 25;11(6):370. doi: 10.3390/toxins11060370. Toxins (Basel). 2019. PMID: 31242582 Free PMC article.
References
-
- Adam-Vizi V., Deri Z., Bors P., Tretter L. Lack of involvement of [Ca2+]i in the external Ca2+-independent release of acetylcholine evoked by veratridine, ouabain and α-latrotoxin: possible role of [Na+]i. J. Physiol. Paris. 1993;87:43–50. - PubMed
-
- Andreev I., Danilevich V.N., Grishin E.V. Alternative splicing of pre-mRNA encoding the Musca domestica latrophilin-like protein (LLP): primary structures of four spliced forms of mRNA and their protein products. Bioorg. Khim. 2005;31:175–185. - PubMed
-
- Ashton A.C., Rahman M.A., Volynski K.E., Manser C., Orlova E.V., Matsushita H., Davletov B.A., van Heel M., Grishin E.V., Ushkaryov Y.A. Tetramerisation of α-latrotoxin by divalent cations is responsible for toxin-induced non-vesicular release and contributes to the Ca2+-dependent vesicular exocytosis from synaptosomes. Biochimie. 2000;82:453–468. - PubMed
-
- Ashton A.C., Volynski K.E., Lelianova V.G., Orlova E.V., Van Renterghem C., Canepari M., Seagar M., Ushkaryov Y.A. α-Latrotoxin, acting via two Ca2+-dependent pathways, triggers exocytosis of two pools of synaptic vesicles. J. Biol. Chem. 2001;276:44695–44703. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
