

Hepatic de novo lipogenesis is present in liver-specific ACC1-deficient mice
- PMID: 17210641
- PMCID: PMC1820479
- DOI: 10.1128/MCB.01122-06
Hepatic de novo lipogenesis is present in liver-specific ACC1-deficient mice
Erratum in
- Mol Cell Biol. 2007 May;27(9):3556
Abstract
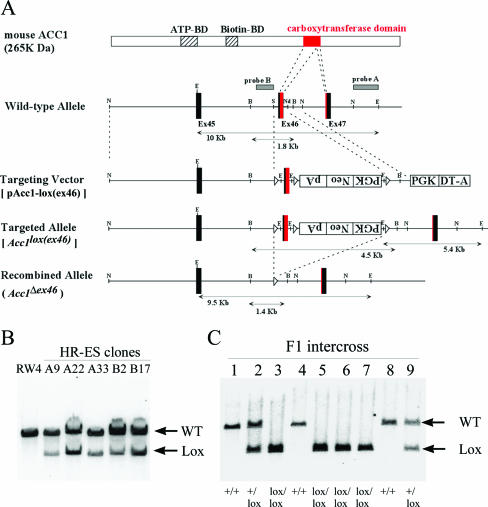
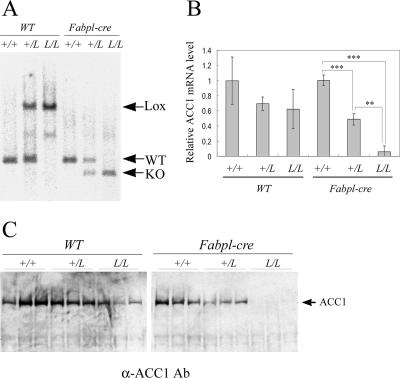
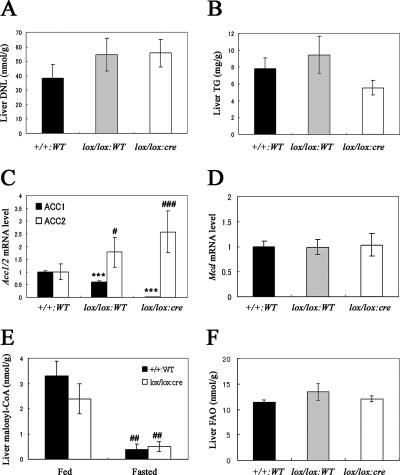
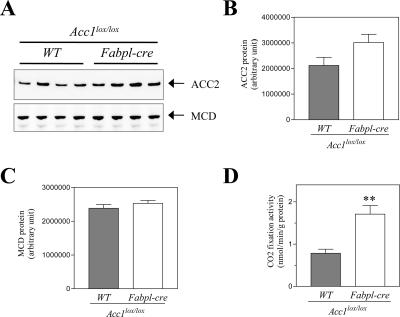
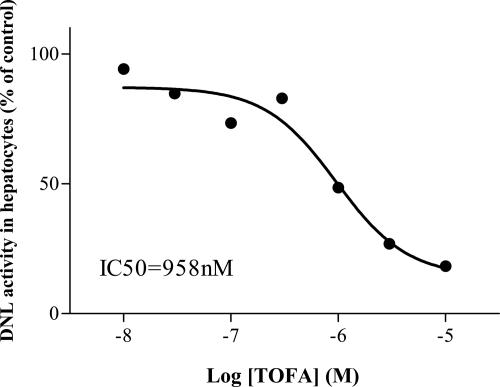
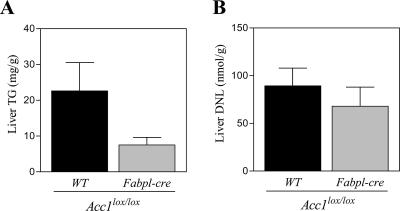
Acetyl coenzyme A (acetyl-CoA) carboxylase (ACC) catalyzes carboxylation of acetyl-CoA to form malonyl-CoA. In mammals, two isozymes exist with distinct physiological roles: cytosolic ACC1 participates in de novo lipogenesis (DNL), and mitochondrial ACC2 is involved in negative regulation of mitochondrial beta-oxidation. Since systemic ACC1 null mice were embryonic lethal, to clarify the physiological role of ACC1 in hepatic DNL, we generated the liver-specific ACC1 null mouse by crossbreeding of an Acc1(lox(ex46)) mouse, in which exon 46 of Acc1 was flanked by two loxP sequences and the liver-specific Cre transgenic mouse. In liver-specific ACC1 null mice, neither hepatic Acc1 mRNA nor protein was detected. However, to compensate for ACC1 function, hepatic ACC2 protein and activity were induced 1.4 and 2.2 times, respectively. Surprisingly, hepatic DNL and malonyl-CoA were maintained at the same physiological levels as in wild-type mice. Furthermore, hepatic DNL was completely inhibited by an ACC1/2 dual inhibitor, 5-tetradecyloxyl-2-furancarboxylic acid. These results strongly demonstrate that malonyl-CoA from ACC2 can access fatty acid synthase and become the substrate for the DNL pathway under the unphysiological circumstances that result with ACC1 disruption. Therefore, there does not appear to be strict compartmentalization of malonyl-CoA from either of the ACC isozymes in the liver.
Figures
References
-
- Abu-Elheiga, L., D. B. Almarza-Ortega, A. Baldini, and S. J. Wakil. 1997. Human acetyl-CoA carboxylase 2. Molecular cloning, characterization, chromosomal mapping, and evidence for two isoforms. J. Biol. Chem. 272:10669-10677. - PubMed
-
- Abu-Elheiga, L., M. M. Matzuk, K. A. Abo-Hashema, and S. J. Wakil. 2001. Continuous fatty acid oxidation and reduced fat storage in mice lacking acetyl-CoA carboxylase 2. Science 291:2613-2616. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous