

Conservation of B class gene expression in the second whorl of a basal grass and outgroups links the origin of lodicules and petals
- PMID: 17210918
- PMCID: PMC1783367
- DOI: 10.1073/pnas.0606434104
Conservation of B class gene expression in the second whorl of a basal grass and outgroups links the origin of lodicules and petals
Abstract
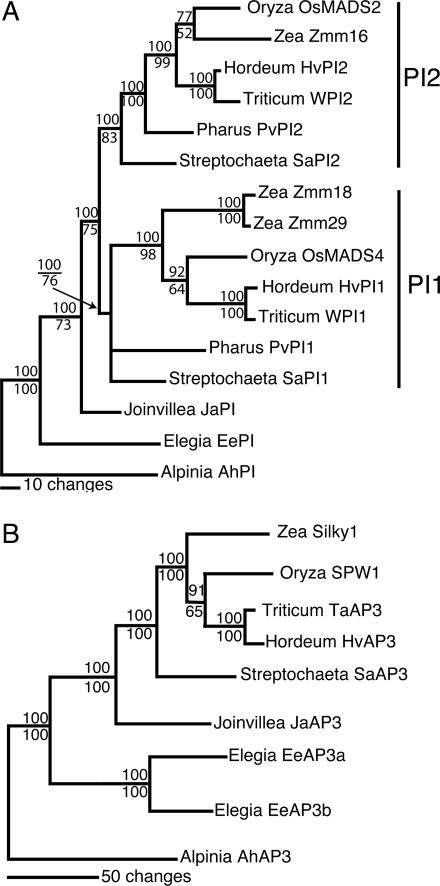
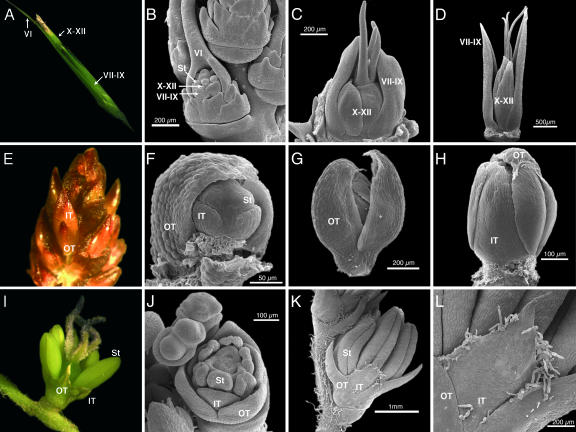
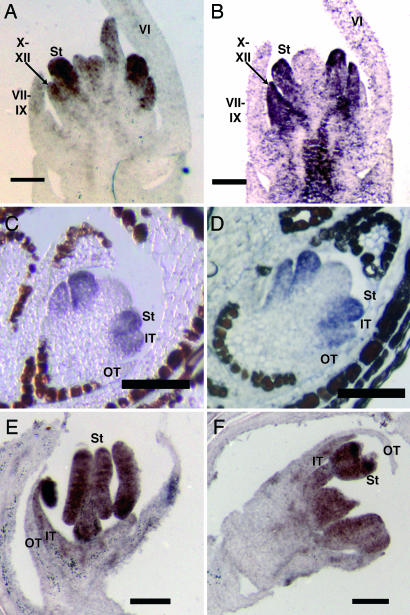
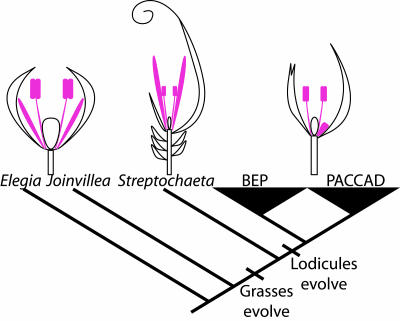
Studies of flower development in core eudicot species have established a central role for B class MADS-box genes in specifying petal and stamen identities. Similarly in maize and rice, B class genes are essential for lodicule and stamen specification, suggesting homology of petals and lodicules and conservation of B class gene activity across angiosperms. However, lodicules are grass-specific organs with a morphology distinct from petals, thus their true homology to eudicot and nongrass monocot floral organs has been a topic of debate. To understand the relationship of lodicules to the sterile floral organs of nongrass monocots we have isolated and observed the expression of B class genes from a basal grass Streptochaeta that diverged before the evolution of lodicules, as well as the outgroups Joinvillea and Elegia, which have a typical monocot floral plan. Our results support a conserved role for B function genes across the angiosperms and provide additional evidence linking the evolution of lodicules and second whorl tepal/petals of monocots. The expression data and morphological analysis suggest that the function of B class genes should be broadly interpreted as required for differentiation of a distinct second floral whorl as opposed to specifying petal identity per se.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Coen ES, Meyerowitz EM. Nature. 1991;353:31–37. - PubMed
-
- Weigel D, Meyerowitz EM. Cell. 1994;78:203–209. - PubMed
-
- Soltis DE, Soltis PS, Albert VA, Oppenheimer DG, dePamphilis CW, Ma H, Frohlich MW, Theissen G. Trends Plants Sci. 2002;7:22–31. discussion 31–34. - PubMed
-
- Mena M, Ambrose BA, Meeley RB, Briggs SP, Yanofsky MF, Schmidt RJ. Science. 1996;274:1537–1540. - PubMed
-
- Ambrose BA, Lerner DR, Ciceri P, Padilla CM, Yanofsky MF, Schmidt RJ. Mol Cell. 2000;5:569–579. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
