

Widespread mRNA polyadenylation events in introns indicate dynamic interplay between polyadenylation and splicing
- PMID: 17210931
- PMCID: PMC1781347
- DOI: 10.1101/gr.5532707
Widespread mRNA polyadenylation events in introns indicate dynamic interplay between polyadenylation and splicing
Abstract
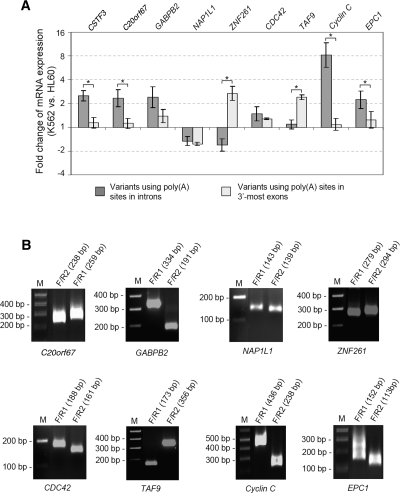
mRNA polyadenylation and pre-mRNA splicing are two essential steps for the maturation of most human mRNAs. Studies have shown that some genes generate mRNA variants involving both alternative polyadenylation and alternative splicing. Polyadenylation in introns can lead to conversion of an internal exon to a 3' terminal exon, which is termed composite terminal exon, or usage of a 3' terminal exon that is otherwise skipped, which is termed skipped terminal exon. Using cDNA/EST and genome sequences, we identified polyadenylation sites in introns for all currently known human genes. We found that approximately 20% human genes have at least one intronic polyadenylation event that can potentially lead to mRNA variants, most of which encode different protein products. The conservation of human intronic poly(A) sites in mouse and rat genomes is lower than that of poly(A) sites in 3'-most exons. Quantitative analysis of a number of mRNA variants generated by intronic poly(A) sites suggests that the intronic polyadenylation activity can vary under different cellular conditions for most genes. Furthermore, we found that weak 5' splice site and large intron size are the determining factors controlling the usage of composite terminal exon poly(A) sites, whereas skipped terminal exon poly(A) sites tend to be associated with strong polyadenylation signals. Thus, our data indicate that dynamic interplay between polyadenylation and splicing leads to widespread polyadenylation in introns and contributes to the complexity of transcriptome in the cell.
Figures




References
-
- Akoulitchev S., Chuikov S., Reinberg D., Chuikov S., Reinberg D., Reinberg D. TFIIH is negatively regulated by cdk8-containing mediator complexes. Nature. 2000;407:102–106. - PubMed
-
- Beaudoing E., Freier S., Wyatt J.R., Claverie J.M., Gautheret D., Freier S., Wyatt J.R., Claverie J.M., Gautheret D., Wyatt J.R., Claverie J.M., Gautheret D., Claverie J.M., Gautheret D., Gautheret D. Patterns of variant polyadenylation signal usage in human genes. Genome Res. 2000;10:1001–1010. - PMC - PubMed
-
- Burge C.B., Tuschl T., Sharp P.A., Tuschl T., Sharp P.A., Sharp P.A. Splicing of precursors to mRNAs by the spliceosomes. In: Gesteland R.F., et al., editors. The RNA world. Cold Spring Harbor Laboratory Press; Cold Spring Harbor, NY: 1999. pp. 525–560.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials