

Requirement of the C3HC4 zinc RING finger of the Arabidopsis PEX10 for photorespiration and leaf peroxisome contact with chloroplasts
- PMID: 17215364
- PMCID: PMC1783365
- DOI: 10.1073/pnas.0610402104
Requirement of the C3HC4 zinc RING finger of the Arabidopsis PEX10 for photorespiration and leaf peroxisome contact with chloroplasts
Abstract
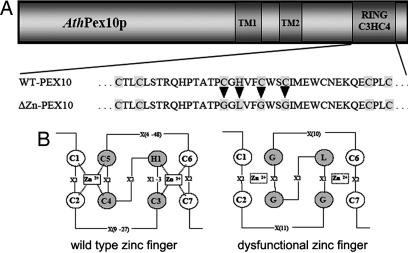
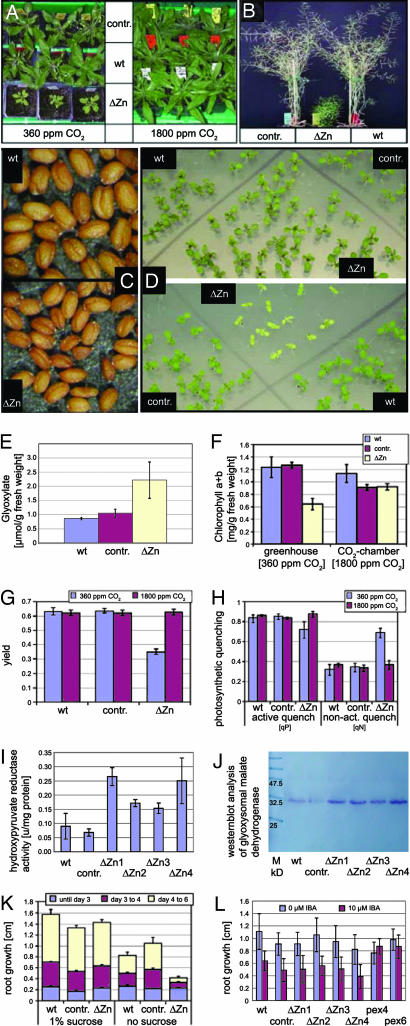
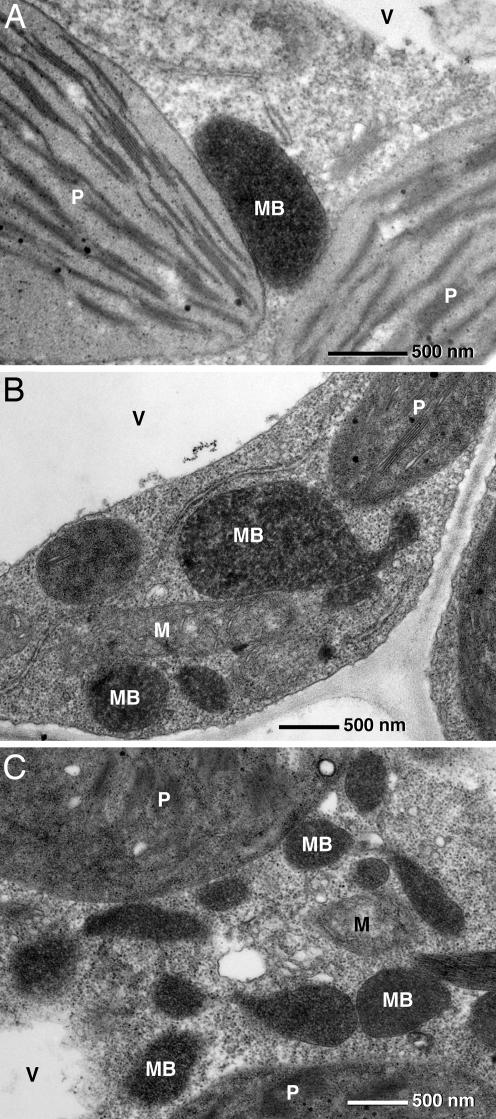
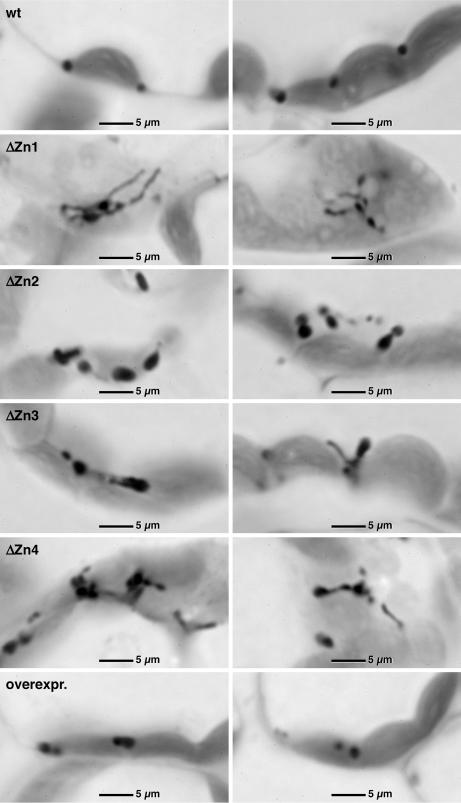
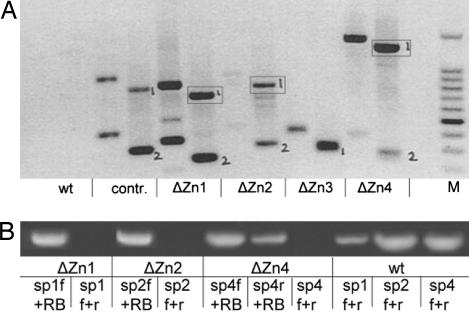
Plant peroxisomes perform multiple vital metabolic processes including lipid mobilization in oil-storing seeds, photorespiration, and hormone biosynthesis. Peroxisome biogenesis requires the function of peroxin (PEX) proteins, including PEX10, a C(3)HC(4) Zn RING finger peroxisomal membrane protein. Loss of function of PEX10 causes embryo lethality at the heart stage. We investigated the function of PEX10 with conditional sublethal mutants. Four T-DNA insertion lines expressing pex10 with a dysfunctional RING finger were created in an Arabidopsis WT background (DeltaZn plants). They could be normalized by growth in an atmosphere of high CO(2) partial pressure, indicating a defect in photorespiration. beta-Oxidation in mutant glyoxysomes was not affected. However, an abnormal accumulation of the photorespiratory metabolite glyoxylate, a lowered content of carotenoids and chlorophyll a and b, and a decreased quantum yield of photosystem II were detected under normal atmosphere, suggesting impaired leaf peroxisomes. Light and transmission electron microscopy demonstrated leaf peroxisomes of the DeltaZn plants to be more numerous, multilobed, clustered, and not appressed to the chloroplast envelope as in WT. We suggest that inactivation of the RING finger domain in PEX10 has eliminated protein interaction required for attachment of peroxisomes to chloroplasts and movement of metabolites between peroxisomes and chloroplasts.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Different functions of the C3HC4 zinc RING finger peroxins PEX10, PEX2, and PEX12 in peroxisome formation and matrix protein import.Proc Natl Acad Sci U S A. 2010 Aug 17;107(33):14915-20. doi: 10.1073/pnas.1009174107. Epub 2010 Aug 2. Proc Natl Acad Sci U S A. 2010. PMID: 20679226 Free PMC article.
-
Peroxisomal ubiquitin-protein ligases peroxin2 and peroxin10 have distinct but synergistic roles in matrix protein import and peroxin5 retrotranslocation in Arabidopsis.Plant Physiol. 2014 Nov;166(3):1329-44. doi: 10.1104/pp.114.247148. Epub 2014 Sep 11. Plant Physiol. 2014. PMID: 25214533 Free PMC article.
-
Genetic Interactions between PEROXIN12 and Other Peroxisome-Associated Ubiquitination Components.Plant Physiol. 2016 Nov;172(3):1643-1656. doi: 10.1104/pp.16.01211. Epub 2016 Sep 20. Plant Physiol. 2016. PMID: 27650450 Free PMC article.
-
Photorespiration: current status and approaches for metabolic engineering.Curr Opin Plant Biol. 2010 Jun;13(3):249-56. doi: 10.1016/j.pbi.2010.01.006. Epub 2010 Feb 23. Curr Opin Plant Biol. 2010. PMID: 20185358 Review.
-
Plant peroxisomes respire in the light: some gaps of the photorespiratory C2 cycle have become filled--others remain.Biochim Biophys Acta. 2006 Dec;1763(12):1496-510. doi: 10.1016/j.bbamcr.2006.09.008. Epub 2006 Sep 14. Biochim Biophys Acta. 2006. PMID: 17046077 Review.
Cited by
-
The ER Is a Common Mediator for the Behavior and Interactions of Other Organelles.Front Plant Sci. 2022 Mar 25;13:846970. doi: 10.3389/fpls.2022.846970. eCollection 2022. Front Plant Sci. 2022. PMID: 35401583 Free PMC article.
-
Dynamics of Peroxisome Homeostasis and Its Role in Stress Response and Signaling in Plants.Front Plant Sci. 2019 Jun 4;10:705. doi: 10.3389/fpls.2019.00705. eCollection 2019. Front Plant Sci. 2019. PMID: 31214223 Free PMC article. Review.
-
Lipid transport required to make lipids of photosynthetic membranes.Photosynth Res. 2018 Dec;138(3):345-360. doi: 10.1007/s11120-018-0545-5. Epub 2018 Jun 30. Photosynth Res. 2018. PMID: 29961189 Review.
-
VpRFP1, a novel C4C4-type RING finger protein gene from Chinese wild Vitis pseudoreticulata, functions as a transcriptional activator in defence response of grapevine.J Exp Bot. 2011 Nov;62(15):5671-82. doi: 10.1093/jxb/err253. Epub 2011 Aug 23. J Exp Bot. 2011. PMID: 21862480 Free PMC article.
-
C2H2-Type Zinc Finger Proteins (DkZF1/2) Synergistically Control Persimmon Fruit Deastringency.Int J Mol Sci. 2019 Nov 9;20(22):5611. doi: 10.3390/ijms20225611. Int J Mol Sci. 2019. PMID: 31717553 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
