

Intestinal epithelial cell-derived interleukin-7: A mechanism for the alteration of intraepithelial lymphocytes in a mouse model of total parenteral nutrition
- PMID: 17215438
- PMCID: PMC1773014
- DOI: 10.1152/ajpgi.00192.2006
Intestinal epithelial cell-derived interleukin-7: A mechanism for the alteration of intraepithelial lymphocytes in a mouse model of total parenteral nutrition
Abstract
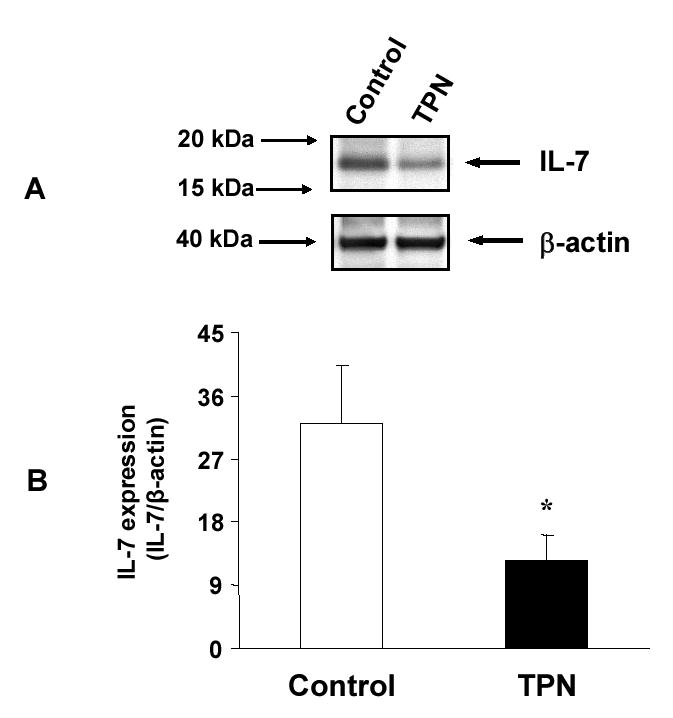
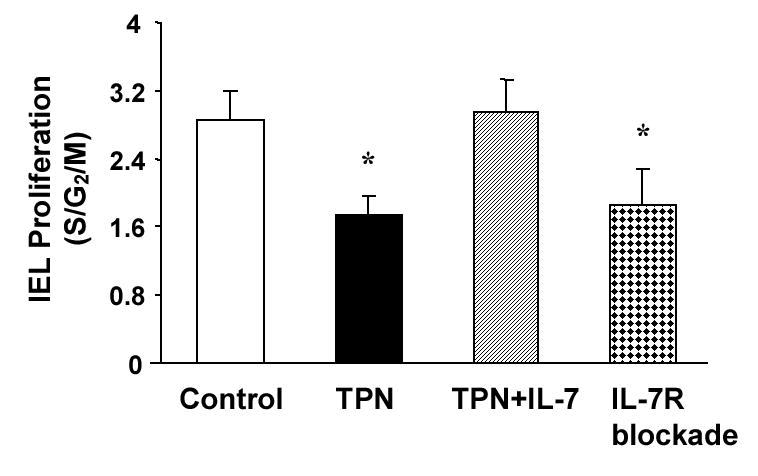
Total parenteral nutrition (TPN), with the absence of enteral nutrition, results in profound changes to both intestinal epithelial cells (EC) as well as the adjacent intraepithelial lymphocyte (IEL) population. Intestinal EC are a rich source of IL-7, a critical factor to support the maintenance of several lymphoid tissues, and TPN results in marked EC changes. On this basis, we hypothesized that TPN would diminish EC-derived IL-7 expression and that this would contribute to the observed changes in the IEL population. Mice received enteral food and intravenous crystalloid solution (control group) or TPN. TPN administration significantly decreased EC-derived IL-7 expression, along with significant changes in IEL phenotype; decreased IEL proliferation; and resulted in a marked decrease in IEL numbers. To better determine the relevance of TPN-related changes in IL-7, TPN mice supplemented with exogenous IL-7 or mice allowed ad libitum feeding and treated with exogenous administration of anti-IL-7 receptor (IL-7R) antibody were also studied. Exogenous IL-7 administration in TPN mice significantly attenuated TPN-associated IEL changes, whereas blocking IL-7R in normal mice resulted in several similar changes in IEL to those observed with TPN. These findings suggest that a decrease in EC-derived IL-7 expression may be a contributing mechanism to account for the observed TPN-associated IEL changes.
Figures






References
-
- Anderson G, Steinberg E. DRG’s and specialized nutritional support: the need for reform. J Paren Enter Nutr. 1986;10:3–10. - PubMed
-
- Faltynek CR, Wang S, Miller D, Young E, Tiberio L, Kross K, Kelley M, Kloszewski E. Administration of human recombinant IL-7 to normal and irradiated mice increases the numbers of lymphocytes and some immature cells of the myeloid lineage. J Immunol. 1992;149:1276–1282. - PubMed
-
- Fujihashi K, Kawabata S, Hiroi T, Yamamoto M, McGhee JR, Nishikawa S, Kiyono H. Interleukin 2 (IL-2) and interleukin 7 (IL-7) reciprocally induce IL-7 and IL-2 receptors on gamma delta T-cell receptor-positive intraepithelial lymphocytes. Proceedings of the National Academy of Sciences of the United States of America. 1996;93:3613–3618. - PMC - PubMed
-
- Fujihashi K, Yamamoto M, McGhee JR, Beagley KW, Kiyono H. Function of alpha beta TCR+ intestinal intraepithelial lymphocytes: Th1- and Th2-type cytokine production by CD4+CD8− and CD4+CD8+ T cells for helper activity. Int Immunol. 1993;5:1473–1481. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials