

EphA4 signaling regulates blastomere adhesion in the Xenopus embryo by recruiting Pak1 to suppress Cdc42 function
- PMID: 17215521
- PMCID: PMC1805096
- DOI: 10.1091/mbc.e06-04-0294
EphA4 signaling regulates blastomere adhesion in the Xenopus embryo by recruiting Pak1 to suppress Cdc42 function
Abstract
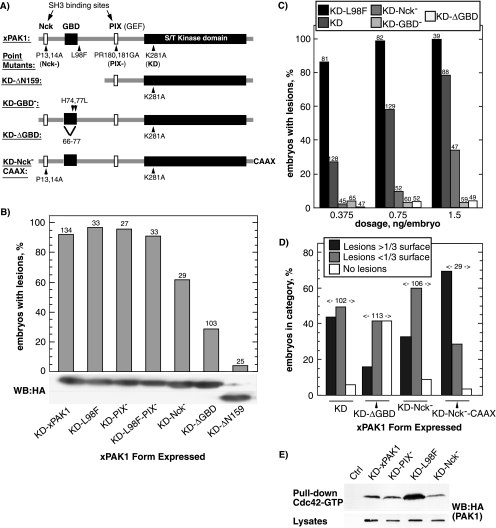
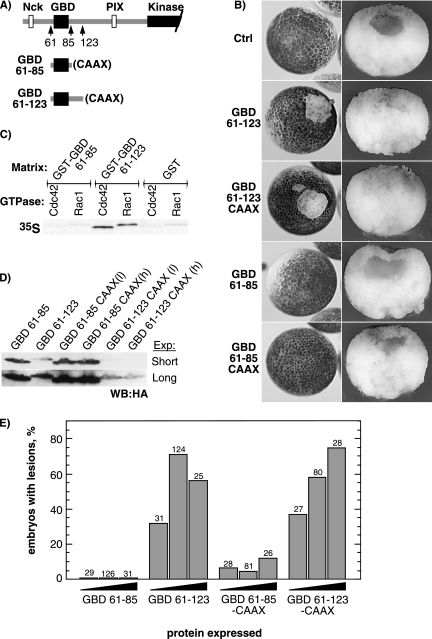
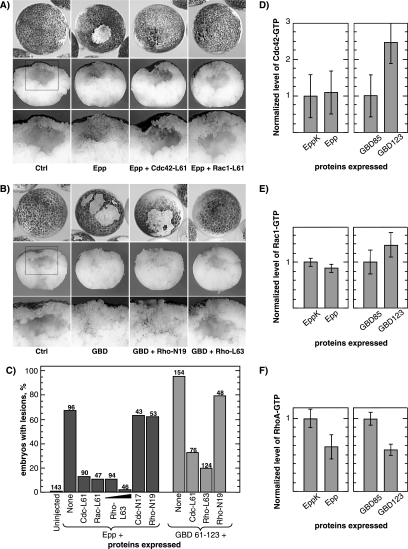
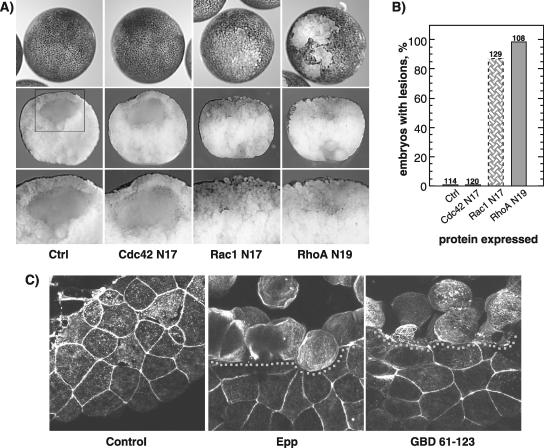
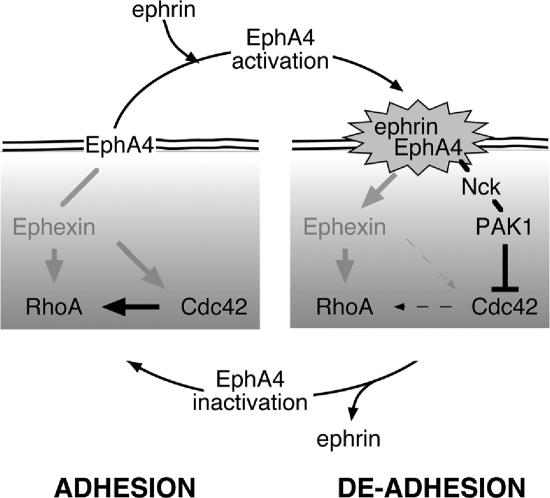
The control of cell adhesion is an important mechanism by which Eph receptors regulate cell sorting during development. Activation of EphA4 in Xenopus blastulae induces a reversible, cell autonomous loss-of-adhesion and disruption of the blastocoel roof. We show this phenotype is rescued by Nckbeta (Grb4) dependent on its interaction with EphA4. Xenopus p21(Cdc42/Rac)-activated kinase xPAK1 interacts with Nck, is activated in embryo by EphA4 in an Nck-dependent manner, and is required for EphA4-induced loss-of-adhesion. Ectopic expression of xPAK1 phenocopies EphA4 activation. This does not require the catalytic activity of xPAK1, but it does require its GTPase binding domain and is enhanced by membrane targeting. Indeed, membrane targeting of the GTPase binding domain (GBD) of xPAK1 alone is sufficient to phenocopy EphA4 loss-of-adhesion. Both EphA4 and the xPAK1-GBD down-regulate RhoA-GTP levels, and consistent with this, loss-of-adhesion can be rescued by activated Cdc42, Rac, and RhoA and can be epistatically induced by dominant-negative RhoA. Despite this, neither Cdc42 nor Rac activities are down-regulated by EphA4 activation or by the xPAK1-GBD. Together, the data suggest that EphA4 activation sequesters active Cdc42 and in this way down-regulates cell-cell adhesion. This novel signaling pathway suggests a mechanism for EphA4-guided migration.
Figures
Similar articles
-
EphA4 catalytic activity causes inhibition of RhoA GTPase in Xenopus laevis embryos.Differentiation. 2002 Mar;70(1):46-55. doi: 10.1046/j.1432-0436.2002.700105.x. Differentiation. 2002. PMID: 11963655
-
Wrch1 is a GTPase-deficient Cdc42-like protein with unusual binding characteristics and cellular effects.Exp Cell Res. 2004 Oct 1;299(2):356-69. doi: 10.1016/j.yexcr.2004.05.029. Exp Cell Res. 2004. PMID: 15350535
-
Activation of cdc42, rac, PAK, and rho-kinase in response to hepatocyte growth factor differentially regulates epithelial cell colony spreading and dissociation.Mol Biol Cell. 2000 May;11(5):1709-25. doi: 10.1091/mbc.11.5.1709. Mol Biol Cell. 2000. PMID: 10793146 Free PMC article.
-
Regulation of phosphorylation pathways by p21 GTPases. The p21 Ras-related Rho subfamily and its role in phosphorylation signalling pathways.Eur J Biochem. 1996 Dec 1;242(2):171-85. doi: 10.1111/j.1432-1033.1996.0171r.x. Eur J Biochem. 1996. PMID: 8973630 Review.
-
p21-activated protein kinase: a crucial component of morphological signaling?Trends Biochem Sci. 1999 Sep;24(9):350-5. doi: 10.1016/s0968-0004(99)01442-5. Trends Biochem Sci. 1999. PMID: 10470034 Review.
Cited by
-
Protein context shapes the specificity of SH3 domain-mediated interactions in vivo.Nat Commun. 2021 Mar 12;12(1):1597. doi: 10.1038/s41467-021-21873-2. Nat Commun. 2021. PMID: 33712617 Free PMC article.
-
A cancer mutation promotes EphA4 oligomerization and signaling by altering the conformation of the SAM domain.J Biol Chem. 2021 Jul;297(1):100876. doi: 10.1016/j.jbc.2021.100876. Epub 2021 Jun 15. J Biol Chem. 2021. PMID: 34139238 Free PMC article.
-
Myosin-X is critical for migratory ability of Xenopus cranial neural crest cells.Dev Biol. 2009 Nov 1;335(1):132-42. doi: 10.1016/j.ydbio.2009.08.018. Epub 2009 Aug 25. Dev Biol. 2009. PMID: 19712673 Free PMC article.
-
Variable combinations of specific ephrin ligand/Eph receptor pairs control embryonic tissue separation.PLoS Biol. 2014 Sep 23;12(9):e1001955. doi: 10.1371/journal.pbio.1001955. eCollection 2014 Sep. PLoS Biol. 2014. PMID: 25247423 Free PMC article.
-
A conserved Oct4/POUV-dependent network links adhesion and migration to progenitor maintenance.Curr Biol. 2013 Nov 18;23(22):2233-2244. doi: 10.1016/j.cub.2013.09.048. Epub 2013 Nov 7. Curr Biol. 2013. PMID: 24210613 Free PMC article.
References
-
- Adler C. E., Miyoshi-Akiyama T., Aleman L. M., Tanaka M., Smith J. M., Mayer B. J. Abl family kinases and Cbl cooperate with the Nck adaptor to modulate Xenopus development. J. Biol. Chem. 2000;275:36472–36478. - PubMed
-
- Bagrodia S., Taylor S. J., Jordon K. A., Van Aelst L., Cerione R. A. A novel regulator of p21-activated kinases. J. Biol. Chem. 1998;273:23633–23636. - PubMed
-
- Benard V., Bohl B. P., Bokoch G. M. Characterization of rac and cdc42 activation in chemoattractant-stimulated human neutrophils using a novel assay for active GTPases. J. Biol. Chem. 1999;274:13198–13204. - PubMed
-
- Bisson N., Islam N., Poitras L., Jean S., Bresnick A. R., Moss T. The catalytic domain of xPAK1 is sufficient to induce myosin II dependent in vivo cell fragmentation independently of other apoptotic events. Dev. Biol. 2003;263:264–281. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
