

Chemical genetics suggests a critical role for lysyl oxidase in zebrafish notochord morphogenesis
- PMID: 17216056
- PMCID: PMC1847539
- DOI: 10.1039/b613673g
Chemical genetics suggests a critical role for lysyl oxidase in zebrafish notochord morphogenesis
Abstract
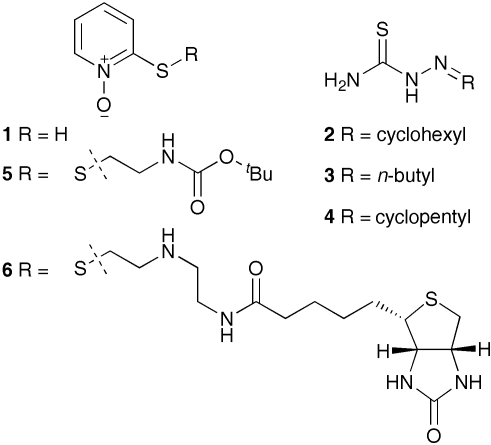
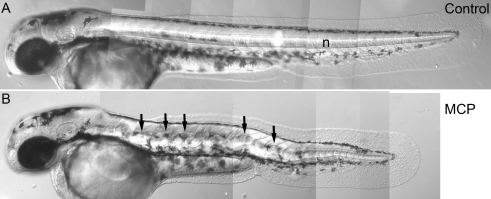
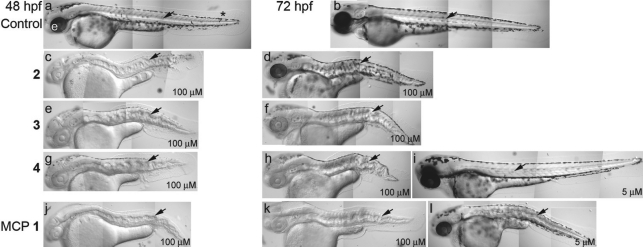
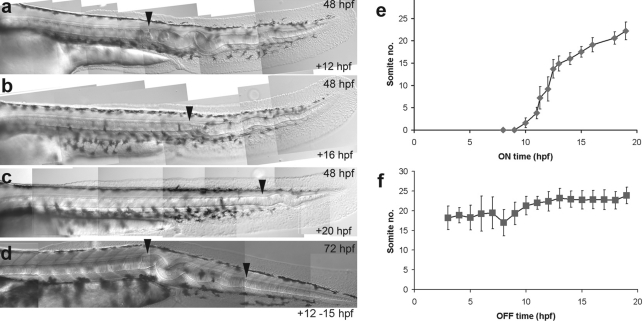
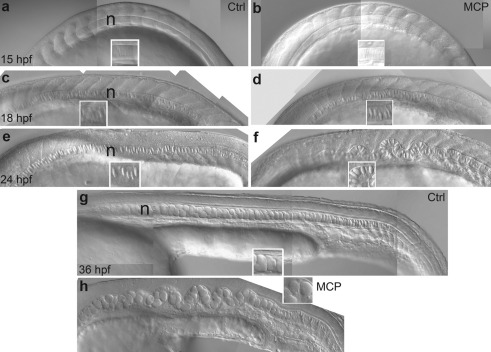
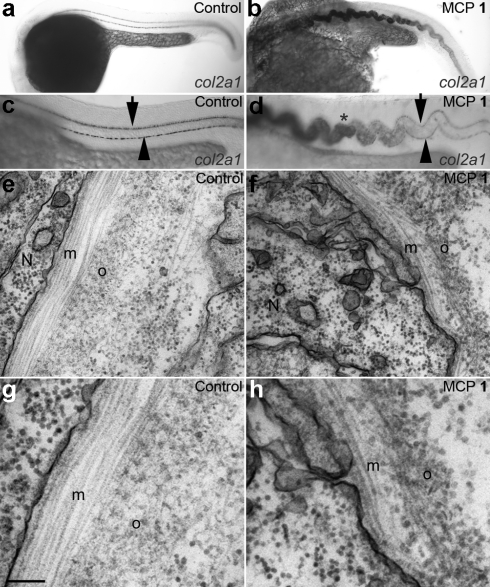
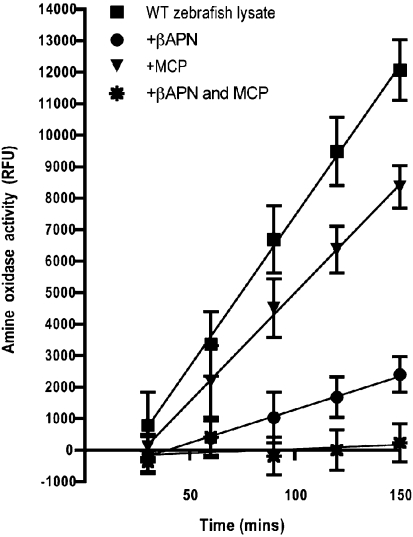
As a result of a chemical genetic screen for modulators of metalloprotease activity, we report that 2-mercaptopyridine-N-oxide induces a conspicuous undulating notochord defect in zebrafish embryos, a phenocopy of the leviathan mutant. The location of the chemically-induced wavy notochord correlated with the timing of application, thus defining a narrow chemical sensitivity window during segmentation stages. Microscopic observations revealed that notochord undulations appeared during the phase of notochord cell vacuolation and notochord elongation. Notochord cells become swollen as well as disorganized, while electron microscopy revealed disrupted organization of collagen fibrils in the surrounding sheath. We demonstrate by assay in zebrafish extracts that 2-mercaptopyridine-N-oxide inhibits lysyl oxidase. Thus, we provide insight into notochord morphogenesis and reveal novel compounds for lysyl oxidase inhibition. Taken together, these data underline the utility of small molecules for elucidating the dynamic mechanisms of early morphogenesis and provide a potential explanation for the recently established role of copper in zebrafish notochord formation.
Figures
References
-
- Spring D. R. Org. Biomol. Chem. 2003;1:3867. - PubMed
-
- Haffter P., Granato M., Brand M., Mullins M. C., Hammerschmidt M., Kane D. A., Odenthal J., van Eeden F. J. M., Jiang Y. J., Heisenberg C. P., Kelsh R. N., Furutani-Seiki M., Vogelsang E., Beuchle D., Schach U., Fabian C., Nusslein-Volhard C. Development. 1996;123:1. - PubMed
-
- Driever W., Solnica-Krezel L., Schier A. F., Neuhauss S. C. F., Malicki J., Stemple D. L., Stainier D. Y. R., Zwartkruis F., Abdelilah S., Rangini Z., Belak J., Boggs C. Development. 1996;123:37. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials