

Ionic currents underlying the response of rat dorsal vagal neurones to hypoglycaemia and chemical anoxia
- PMID: 17218356
- PMCID: PMC2151378
- DOI: 10.1113/jphysiol.2006.126094
Ionic currents underlying the response of rat dorsal vagal neurones to hypoglycaemia and chemical anoxia
Abstract
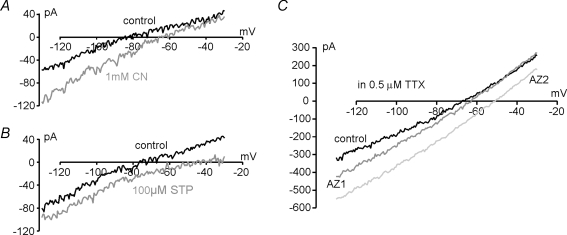
A proportion of dorsal vagal neurones (DVN) are glucosensors. These cells respond to brief hypoglycaemia either with a K(ATP) channel-mediated hyperpolarization or with depolarization owing to an as yet unknown mechanism. K(ATP) currents are observed not only during hypoglycaemia, but also in response to mitochondrial inhibition. Here we show that similarly to the observations for K(ATP) currents, both hypoglycaemia and inhibition of mitochondrial function elicited a small inward current that persisted in TTX in DVN of rat brainstem slices. Removal of glucose from the bath solution induced this inward current within 50 +/- 4 s in one subpopulation of DVN and in 279 +/- 36 s in another subpopulation. No such subpopulations were observed for the response to mitochondrial inhibition. Biophysical analysis revealed that mitochondrial inhibition or hypoglycaemia inhibited an openly rectifying K+ conductance in 25% of DVN. In the remaining cells, either an increase in conductance, with a reversal potential between -58 and +10 mV, or a parallel inward shift of the holding current was observed. This current most probably resulted from inhibition of the Na+-K+-ATPase and/or the opening of an ion channel. Recordings with electrodes containing 145 mm instead of 5 mm Cl- failed to shift the reversal potential of the inward current, indicating that a Cl- channel was not involved. In summary, glucosensing and non-glucosensing DVN appear to use common electrical pathways to respond to mitochondrial inhibition and to hypoglycaemia. We suggest that differences in glucose metabolism rather than differences in the complement of ion channels distinguish these two cell types.
Figures





References
-
- Ashcroft FM. Adenosine 5′-triphosphate-sensitive potassium channels. Annu Rev Neurosci. 1988;11:97–118. - PubMed
-
- Ashford ML, Boden PR, Treherne JM. Glucose-induced excitation of hypothalamic neurones is mediated by ATP-sensitive K+ channels. Pflugers Arch. 1990;415:479–483. - PubMed
-
- Ballanyi K, Kulik A. Intracellular Ca2+ during metabolic activation of KATP channels in spontaneously active dorsal vagal neurons in medullary slices. Eur J Neurosci. 1998;10:2574–2585. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical