

DELLAs contribute to plant photomorphogenesis
- PMID: 17220364
- PMCID: PMC1820925
- DOI: 10.1104/pp.106.092254
DELLAs contribute to plant photomorphogenesis
Abstract
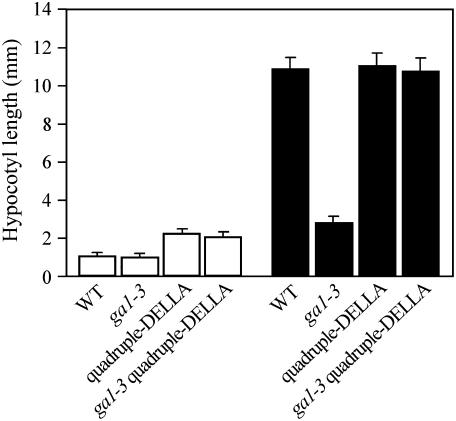
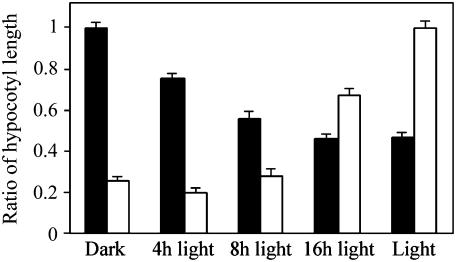
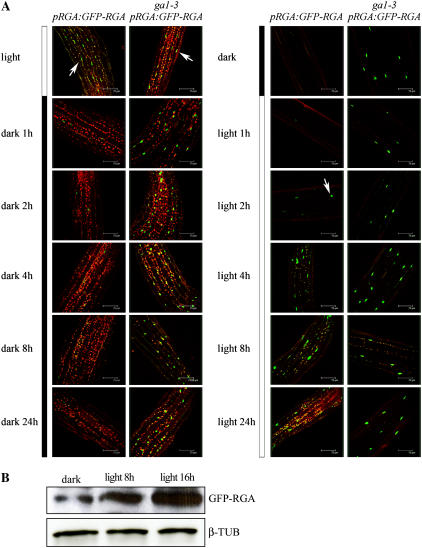
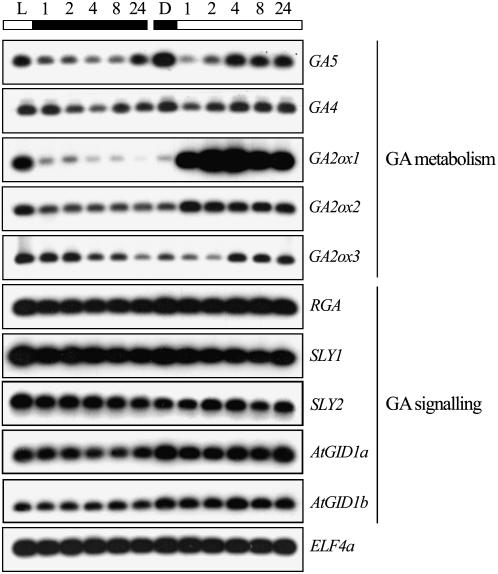
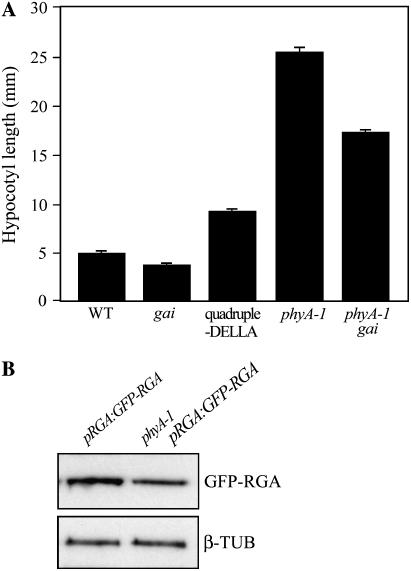
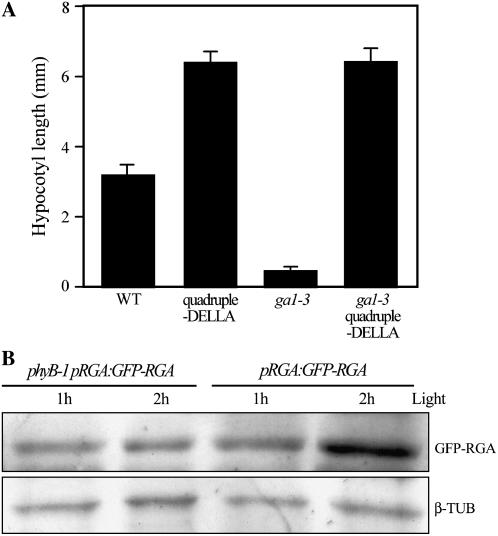
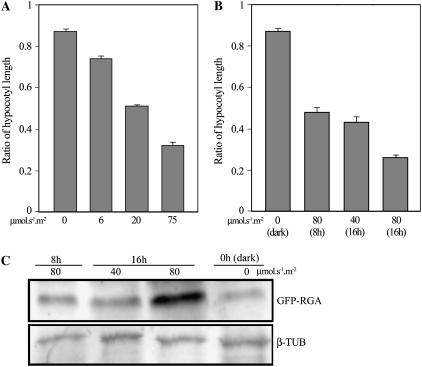
Plant morphogenesis is profoundly influenced by light (a phenomenon known as photomorphogenesis). For example, light inhibits seedling hypocotyl growth via activation of phytochromes and additional photoreceptors. Subsequently, information is transmitted through photoreceptor-linked signal transduction pathways and used (via previously unknown mechanisms) to control hypocotyl growth. Here we show that light inhibition of Arabidopsis (Arabidopsis thaliana) hypocotyl growth is in part dependent on the DELLAs (a family of nuclear growth-restraining proteins that mediate the effect of the phytohormone gibberellin [GA] on growth). We show that light inhibition of growth is reduced in DELLA-deficient mutant hypocotyls. We also show that light activation of phytochromes promotes the accumulation of DELLAs. A green fluorescent protein (GFP)-tagged DELLA (GFP-RGA) accumulates in elongating cells of light-grown, but not dark-grown, transgenic wild-type hypocotyls. Furthermore, transfer of seedlings from light to dark (or vice versa) results in rapid changes in hypocotyl GFP-RGA accumulation, changes that are paralleled by rapid alterations in the abundance in hypocotyls of transcripts encoding enzymes of GA metabolism. These observations suggest that light-dependent changes in hypocotyl GFP-RGA accumulation are a consequence of light-dependent changes in bioactive GA level. Finally, we show that GFP accumulation and quantitative modulation of hypocotyl growth is proportionate with light energy dose (the product of exposure duration and fluence rate). Hence, DELLAs inhibit hypocotyl growth during the light phase of the day-night cycle via a mechanism that is quantitatively responsive to natural light variability. We conclude that DELLAs are a major component of the adaptively significant mechanism via which light regulates plant growth during photomorphogenesis.
Figures
References
-
- Achard P, Cheng H, De Grauwe L, Decat J, Schoutteten H, Moritz T, Van Der Straeten D, Peng J, Harberd NP (2006) Integration of plant responses to environmentally activated phytohormonal signals. Science 331 91–94 - PubMed
-
- Alvey L, Harberd NP (2005) DELLA proteins: integrators of multiple plant growth regulatory inputs? Physiol Plant 123 153–160
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
