

Increased genome instability in Escherichia coli lon mutants: relation to emergence of multiple-antibiotic-resistant (Mar) mutants caused by insertion sequence elements and large tandem genomic amplifications
- PMID: 17220404
- PMCID: PMC1855481
- DOI: 10.1128/AAC.01128-06
Increased genome instability in Escherichia coli lon mutants: relation to emergence of multiple-antibiotic-resistant (Mar) mutants caused by insertion sequence elements and large tandem genomic amplifications
Abstract
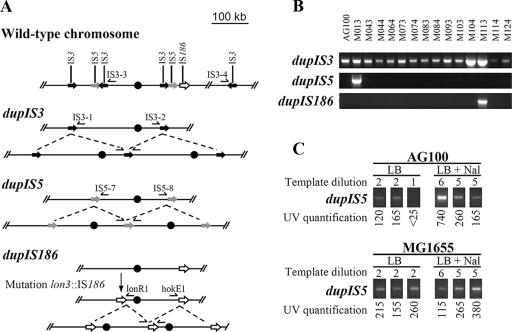
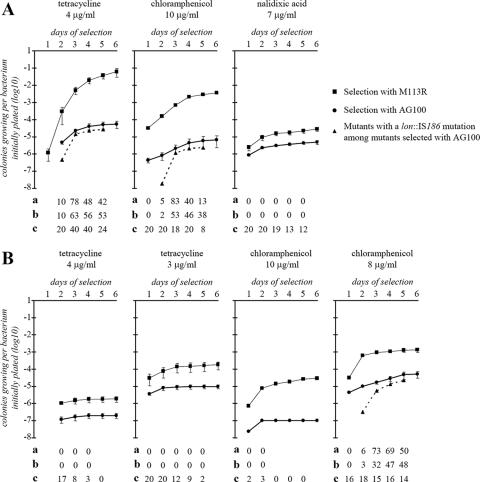
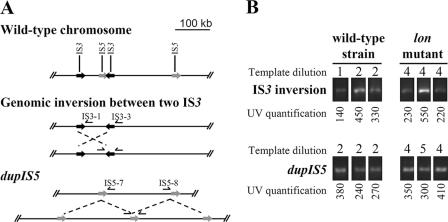
Thirteen spontaneous multiple-antibiotic-resistant (Mar) mutants of Escherichia coli AG100 were isolated on Luria-Bertani (LB) agar in the presence of tetracycline (4 microg/ml). The phenotype was linked to insertion sequence (IS) insertions in marR or acrR or unstable large tandem genomic amplifications which included acrAB and which were bordered by IS3 or IS5 sequences. Five different lon mutations, not related to the Mar phenotype, were also found in 12 of the 13 mutants. Under specific selective conditions, most drug-resistant mutants appearing late on the selective plates evolved from a subpopulation of AG100 with lon mutations. That the lon locus was involved in the evolution to low levels of multidrug resistance was supported by the following findings: (i) AG100 grown in LB broth had an important spontaneous subpopulation (about 3.7x10(-4)) of lon::IS186 mutants, (ii) new lon mutants appeared during the selection on antibiotic-containing agar plates, (iii) lon mutants could slowly grow in the presence of low amounts (about 2x MIC of the wild type) of chloramphenicol or tetracycline, and (iv) a lon mutation conferred a mutator phenotype which increased IS transposition and genome rearrangements. The association between lon mutations and mutations causing the Mar phenotype was dependent on the medium (LB versus MacConkey medium) and the antibiotic used for the selection. A previously reported unstable amplifiable high-level resistance observed after the prolonged growth of Mar mutants in a low concentration of tetracycline or chloramphenicol can be explained by genomic amplification.
Figures
Similar articles
-
Role for tandem duplication and lon protease in AcrAB-TolC- dependent multiple antibiotic resistance (Mar) in an Escherichia coli mutant without mutations in marRAB or acrRAB.J Bacteriol. 2006 Jun;188(12):4413-23. doi: 10.1128/JB.01502-05. J Bacteriol. 2006. PMID: 16740948 Free PMC article.
-
Lon protease inactivation, or translocation of the lon gene, potentiate bacterial evolution to antibiotic resistance.Mol Microbiol. 2013 Dec;90(6):1233-48. doi: 10.1111/mmi.12429. Epub 2013 Oct 30. Mol Microbiol. 2013. PMID: 24325250
-
Roles of Lon protease and its substrate MarA during sodium salicylate-mediated growth reduction and antibiotic resistance in Escherichia coli.Microbiology (Reading). 2016 May;162(5):764-776. doi: 10.1099/mic.0.000271. Epub 2016 Mar 4. Microbiology (Reading). 2016. PMID: 26944926
-
The multiple antibiotic resistance (mar) locus and its significance.Res Vet Sci. 2002 Apr;72(2):87-93. doi: 10.1053/rvsc.2001.0537. Res Vet Sci. 2002. PMID: 12027588 Review.
-
[Analysis of Drug Resistance Using Experimental Evolution].Yakugaku Zasshi. 2017;137(4):373-376. doi: 10.1248/yakushi.16-00235-1. Yakugaku Zasshi. 2017. PMID: 28381708 Review. Japanese.
Cited by
-
The FlyCatwalk: a high-throughput feature-based sorting system for artificial selection in Drosophila.G3 (Bethesda). 2015 Jan 2;5(3):317-27. doi: 10.1534/g3.114.013664. G3 (Bethesda). 2015. PMID: 25556112 Free PMC article.
-
Identification of Evolutionary Trajectories Associated with Antimicrobial Resistance Using Microfluidics.ACS Infect Dis. 2022 Jan 14;8(1):242-254. doi: 10.1021/acsinfecdis.1c00564. Epub 2021 Dec 28. ACS Infect Dis. 2022. PMID: 34962128 Free PMC article.
-
IS1-mediated chromosomal amplification of the arn operon leads to polymyxin B resistance in Escherichia coli B strains.mBio. 2024 Jul 17;15(7):e0063424. doi: 10.1128/mbio.00634-24. Epub 2024 Jun 21. mBio. 2024. PMID: 38904391 Free PMC article.
-
The challenge of efflux-mediated antibiotic resistance in Gram-negative bacteria.Clin Microbiol Rev. 2015 Apr;28(2):337-418. doi: 10.1128/CMR.00117-14. Clin Microbiol Rev. 2015. PMID: 25788514 Free PMC article. Review.
-
Proteostasis modulates gene dosage evolution in antibiotic-resistant bacteria.Elife. 2025 Mar 12;13:RP99785. doi: 10.7554/eLife.99785. Elife. 2025. PMID: 40073078 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
