

Repeated recruitment of LTR retrotransposons as promoters by the anti-apoptotic locus NAIP during mammalian evolution
- PMID: 17222062
- PMCID: PMC1781489
- DOI: 10.1371/journal.pgen.0030010
Repeated recruitment of LTR retrotransposons as promoters by the anti-apoptotic locus NAIP during mammalian evolution
Abstract
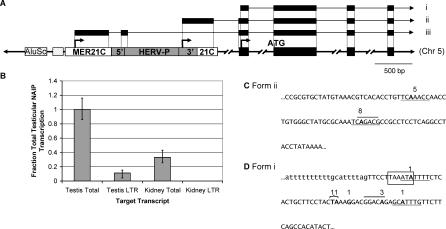
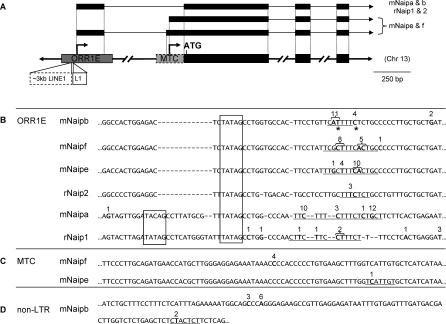
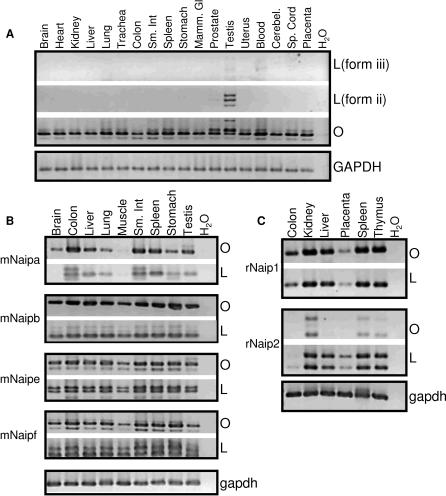
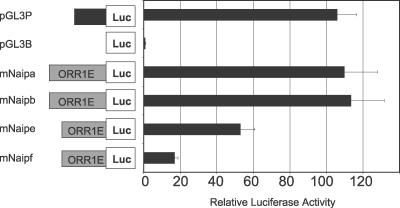
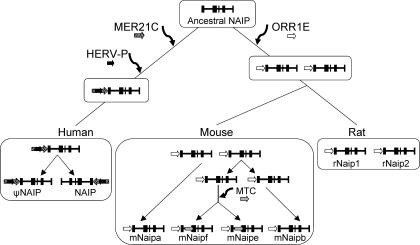
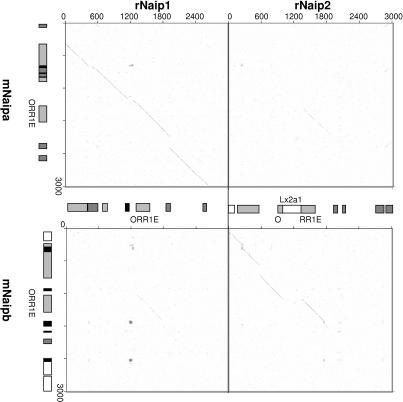
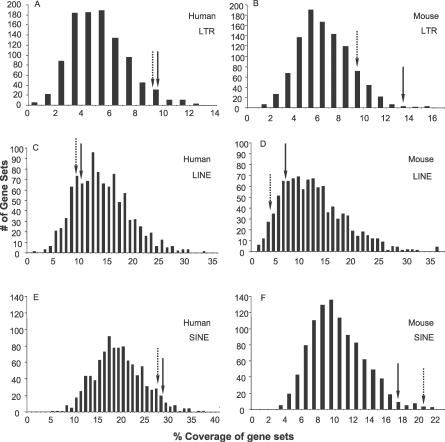
Neuronal apoptosis inhibitory protein (NAIP, also known as BIRC1) is a member of the conserved inhibitor of apoptosis protein (IAP) family. Lineage-specific rearrangements and expansions of this locus have yielded different copy numbers among primates and rodents, with human retaining a single functional copy and mouse possessing several copies, depending on the strain. Roles for this gene in disease have been documented, but little is known about transcriptional regulation of NAIP. We show here that NAIP has multiple promoters sharing no similarity between human and rodents. Moreover, we demonstrate that multiple, domesticated long terminal repeats (LTRs) of endogenous retroviral elements provide NAIP promoter function in human, mouse, and rat. In human, an LTR serves as a tissue-specific promoter, active primarily in testis. However, in rodents, our evidence indicates that an ancestral LTR common to all rodent genes is the major, constitutive promoter for these genes, and that a second LTR found in two of the mouse genes is a minor promoter. Thus, independently acquired LTRs have assumed regulatory roles for orthologous genes, a remarkable evolutionary scenario. We also demonstrate that 5' flanking regions of IAP family genes as a group, in both human and mouse are enriched for LTR insertions compared to average genes. We propose several potential explanations for these findings, including a hypothesis that recruitment of LTRs near NAIP or other IAP genes may represent a host-cell adaptation to modulate apoptotic responses.
Conflict of interest statement
Competing interests. The authors have declared that no competing interests exist.
Figures
References
-
- Lander ES, Linton LM, Birren B, Nusbaum C, Zody MC, et al. Initial sequencing and analysis of the human genome. Nature. 2001;409:860–921. - PubMed
-
- Waterston RH, Lindblad-Toh K, Birney E, Rogers J, Abril JF, et al. Initial sequencing and comparative analysis of the mouse genome. Nature. 2002;420:520–562. - PubMed
-
- Nekrutenko A, Li WH. Transposable elements are found in a large number of human protein-coding genes. Trends Genet. 2001;17:619–621. - PubMed
-
- Gotea V, Makalowski W. Do transposable elements really contribute to proteomes? Trends Genet. 2006;22:260–267. - PubMed
-
- Jordan IK, Rogozin IB, Glazko GV, Koonin EV. Origin of a substantial fraction of human regulatory sequences from transposable elements. Trends Genet. 2003;19:68–72. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
