

High mobility group A2 is a target for miRNA-98 in head and neck squamous cell carcinoma
- PMID: 17222355
- PMCID: PMC1783857
- DOI: 10.1186/1476-4598-6-5
High mobility group A2 is a target for miRNA-98 in head and neck squamous cell carcinoma
Abstract
Background: HMGA2 expression has been shown to be associated with enhanced selective chemosensitivity towards the topoisomerase (topo) II inhibitor, doxorubicin, in cancer cells. Although the roles of signaling cascades and proteins as regulatory factors in development, neoplasia and adaptation to the environment are becoming well established, evidence for the involvement of regulatory small RNA molecules, such as microRNAs (miRNAs) as important regulators of both transcriptional and posttranscriptional gene silencing is presently mounting.
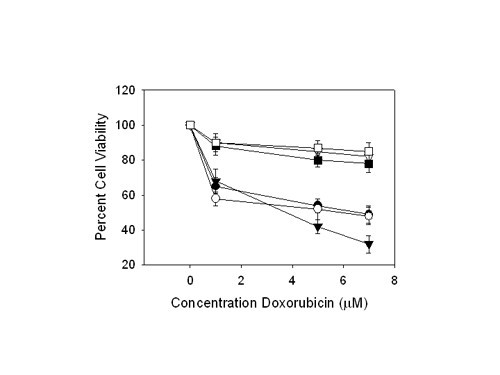
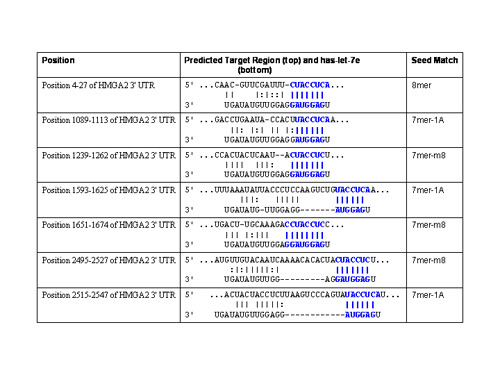
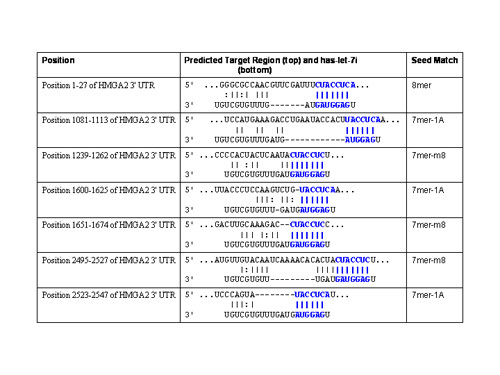
Results: Here we report that HMGA2 expression in head and neck squamous cell carcinoma (HNSCC) cells is regulated in part by miRNA-98 (miR-98). Albeit HMGA2 is associated with enhanced selective chemosensitivity towards topoisomerase (topo) II inhibitor, doxorubicin in HNSCC, the expression of HMGA2 is thwarted by hypoxia. This is accompanied by enhanced expression of miRNA-98 and other miRNAs, which predictably target HMGA2. Moreover, we show that transfection of pre-miR-98trade mark during normoxia diminishes HMGA2 and potentiates resistance to doxorubicin and cisplatin. These findings implicate the role of a miRNA as a key element in modulating tumors in variable microenvironments.
Conclusion: These studies validate the observation that HMGA2 plays a prominent role in governing genotoxic responses. However, this may only represent cells growing under normal oxygen tensions. The demonstration that miRNA profiles are altered during hypoxia and repress a genotoxic response indicates that changes in microenvironment in eukaryotes mimic those of lower species and plants, where, for example, abiotic stresses regulate the expression of thousands of genes in plants at both transcriptional and posttranscriptional levels through a number of miRNAs and other small regulatory RNAs.
Figures






References
-
- Boo LM, Lin HH, Chung V, Zhou B, Louie SG, O'Reilly MA, Yen Y, Ann DK. High mobility group A2 potentiates genotoxic stress in part through the modulation of basal and DNA damage-dependent phosphatidylinositol 3-kinase-related protein kinase activation. Cancer Res. 2005;65:6622–6630. doi: 10.1158/0008-5472.CAN-05-0086. - DOI - PubMed
-
- Reeves R, Beckerbauer L. HMGI/Y proteins: flexible regulators of transcription and chromatin structure. Biochim Biophys Acta. 2001;1519:13–29. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
