

Periostin regulates collagen fibrillogenesis and the biomechanical properties of connective tissues
- PMID: 17226767
- PMCID: PMC3393091
- DOI: 10.1002/jcb.21224
Periostin regulates collagen fibrillogenesis and the biomechanical properties of connective tissues
Abstract
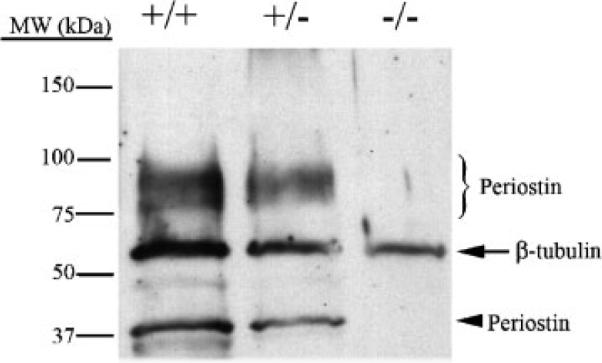
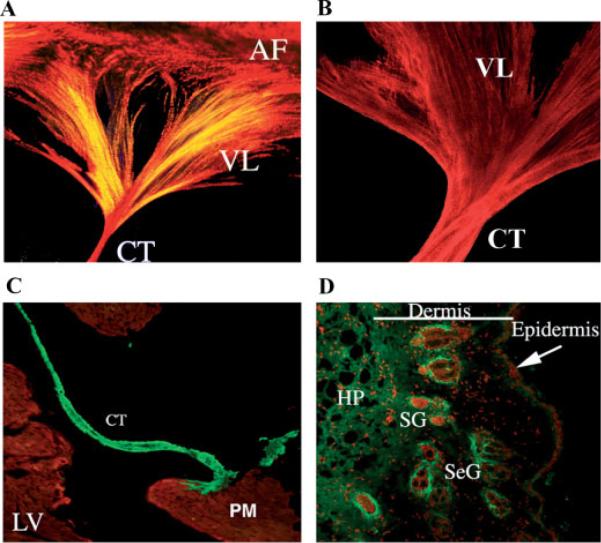
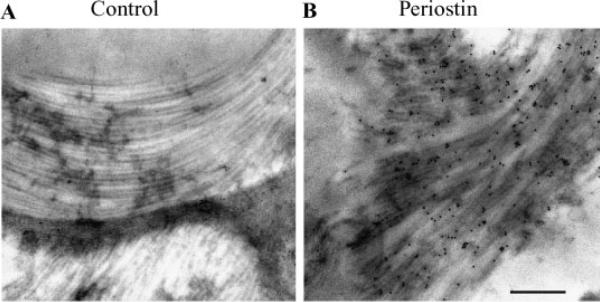
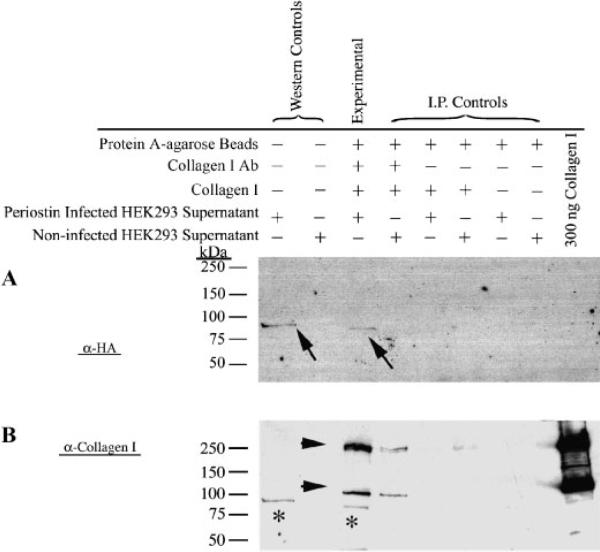
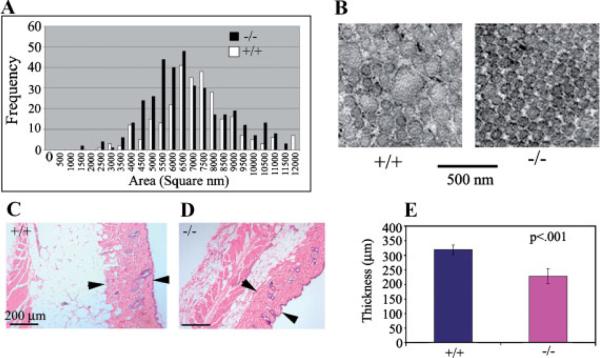
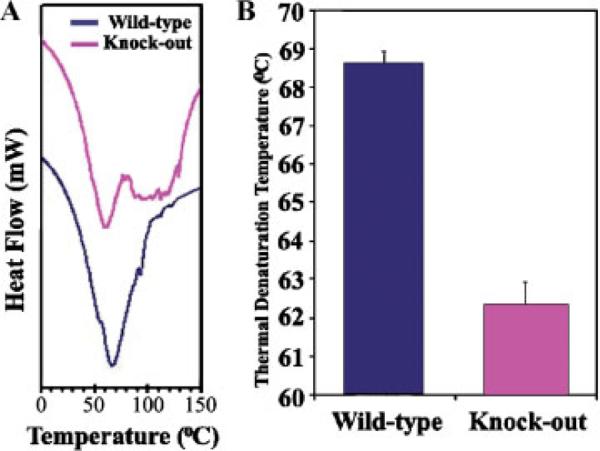
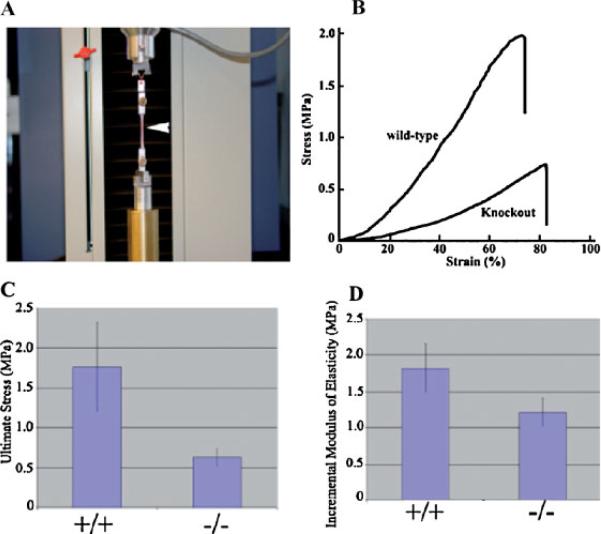
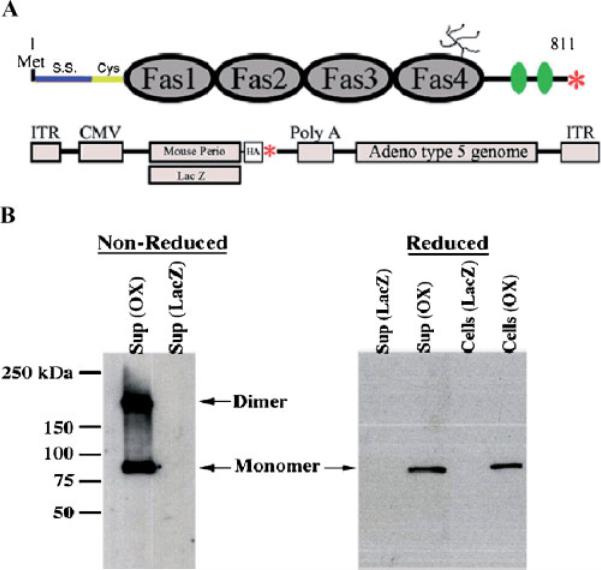
Periostin is predominantly expressed in collagen-rich fibrous connective tissues that are subjected to constant mechanical stresses including: heart valves, tendons, perichondrium, cornea, and the periodontal ligament (PDL). Based on these data we hypothesize that periostin can regulate collagen I fibrillogenesis and thereby affect the biomechanical properties of connective tissues. Immunoprecipitation and immunogold transmission electron microscopy experiments demonstrate that periostin is capable of directly interacting with collagen I. To analyze the potential role of periostin in collagen I fibrillogenesis, gene targeted mice were generated. Transmission electron microscopy and morphometric analyses demonstrated reduced collagen fibril diameters in skin dermis of periostin knockout mice, an indication of aberrant collagen I fibrillogenesis. In addition, differential scanning calorimetry (DSC) demonstrated a lower collagen denaturing temperature in periostin knockout mice, reflecting a reduced level of collagen cross-linking. Functional biomechanical properties of periostin null skin specimens and atrioventricular (AV) valve explant experiments provided direct evidence of the role that periostin plays in regulating the viscoelastic properties of connective tissues. Collectively, these data demonstrate for the first time that periostin can regulate collagen I fibrillogenesis and thereby serves as an important mediator of the biomechanical properties of fibrous connective tissues.
Figures
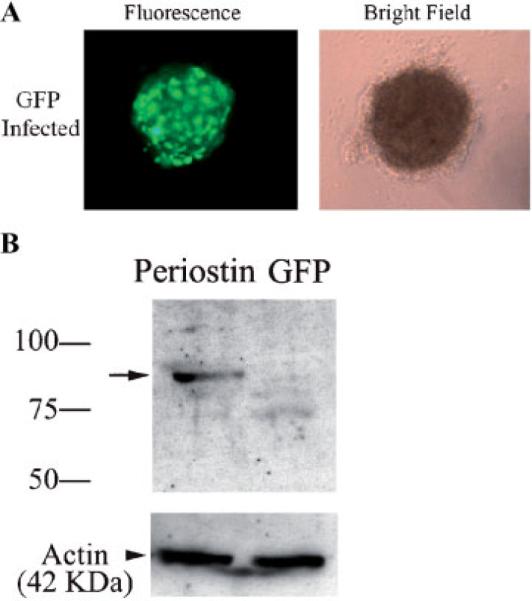
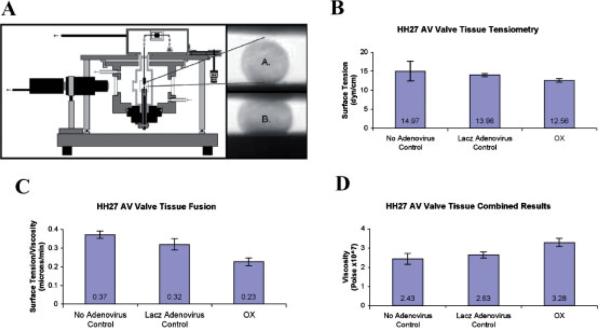
References
-
- Ameye L, Young MF. Mice deficient in small leucine-rich proteoglycans: Novel in vivo models for osteoporosis, osteoarthritis, Ehlers-Danlos syndrome, muscular dystrophy, and corneal diseases. Glycobiology. 2002;12:107R–116R. - PubMed
-
- Ameye L, Aria D, Jepsen K, Oldberg A, Xu T, Young MF. Abnormal collagen fibrils in tendons of biglycan/ fibromodulin-deficient mice lead to gait impairment, ectopic ossification, and osteoarthritis. FASEB J. 2002;16:673–680. - PubMed
-
- Beck GR, Jr., Moran E, Knecht N. Inorganic phosphate regulates multiple genes during osteoblast differentiation, including Nrf2. Exp Cell Res. 2003;288:288–300. - PubMed
-
- Berisio R, Vitagliano L, Mazzarella L, Zagari A. Recent progress on collagen triple helix structure, stability and assembly. Protein Pept Lett. 2002;9:107–116. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- HL52813/HL/NHLBI NIH HHS/United States
- R01 HL066231/HL/NHLBI NIH HHS/United States
- R01 HL072958/HL/NHLBI NIH HHS/United States
- P20RR016434/RR/NCRR NIH HHS/United States
- 5MO1RR001070-28/RR/NCRR NIH HHS/United States
- R01 HL033756/HL/NHLBI NIH HHS/United States
- R01 HL077342/HL/NHLBI NIH HHS/United States
- R01 HL33756/HL/NHLBI NIH HHS/United States
- R01 HL66231/HL/NHLBI NIH HHS/United States
- EF0526854/PHS HHS/United States
- P20 RR016434/RR/NCRR NIH HHS/United States
- P01 HL052813/HL/NHLBI NIH HHS/United States
- R01 HL060714/HL/NHLBI NIH HHS/United States
- M01 RR001070/RR/NCRR NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
