

Breast cancer metastasis suppressor 1 (BRMS1) inhibits osteopontin transcription by abrogating NF-kappaB activation
- PMID: 17227585
- PMCID: PMC1796551
- DOI: 10.1186/1476-4598-6-6
Breast cancer metastasis suppressor 1 (BRMS1) inhibits osteopontin transcription by abrogating NF-kappaB activation
Abstract
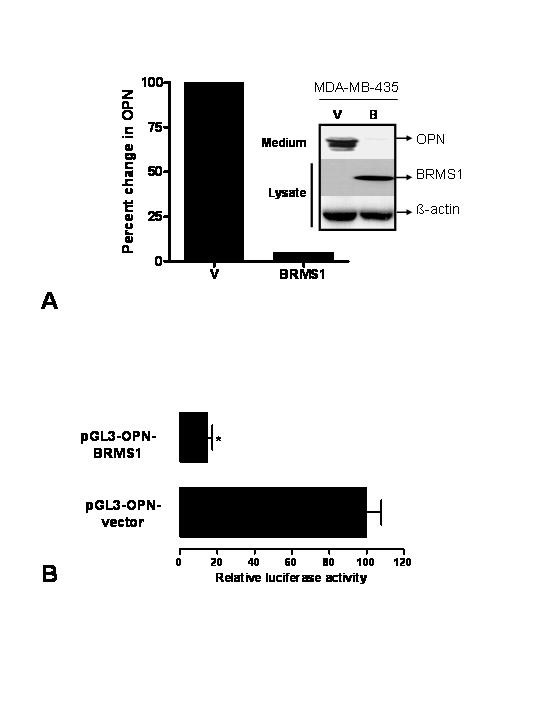
Background: Osteopontin (OPN), a secreted phosphoglycoprotein, has been strongly associated with tumor progression and aggressive cancers. MDA-MB-435 cells secrete very high levels of OPN. However metastasis-suppressed MDA-MB-435 cells, which were transfected with breast cancer metastasis suppressor 1 (BRMS1), expressed significantly less OPN. BRMS1 is a member of mSin3-HDAC transcription co-repressor complex and has been shown to suppress the metastasis of breast cancer and melanoma cells in animal models. Hence we hypothesized that BRMS1 regulates OPN expression.
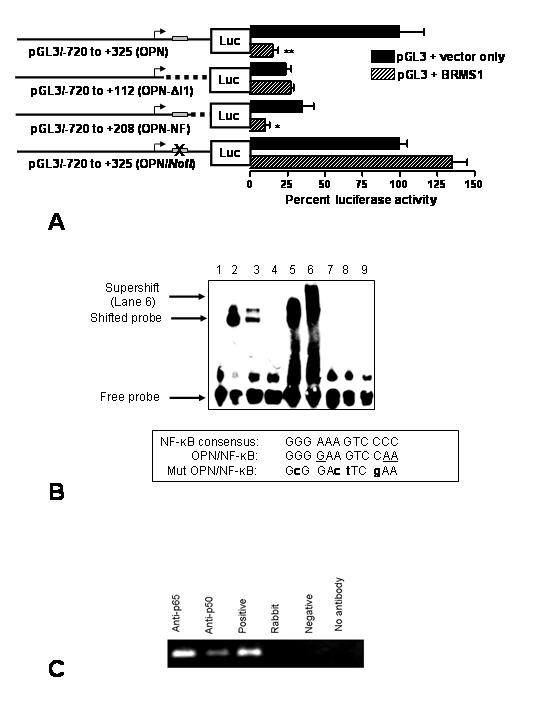
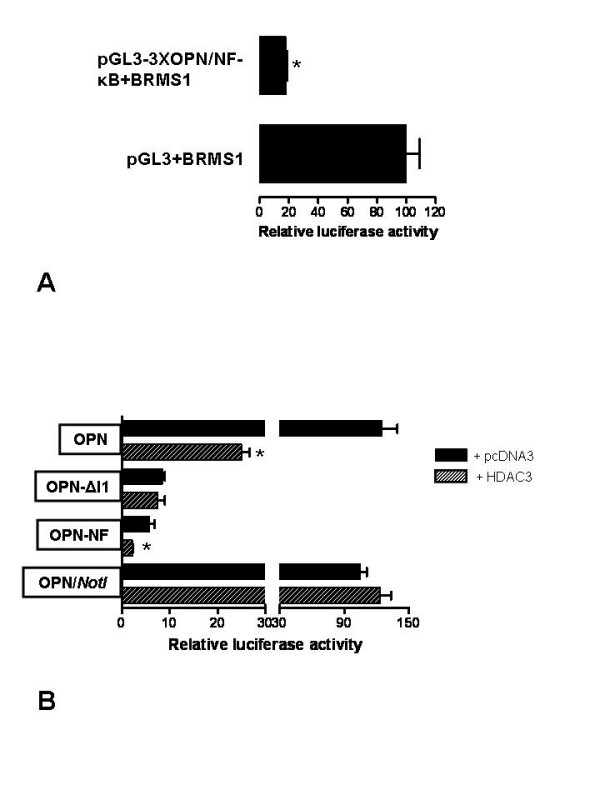
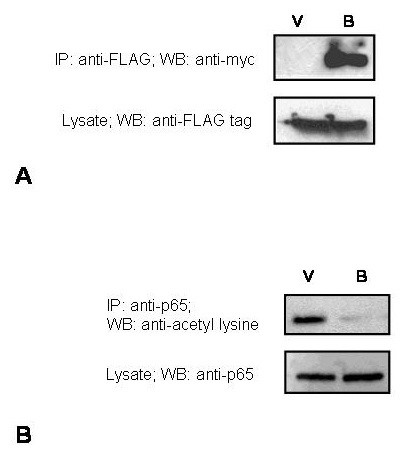
Results: The search for a BRMS1-regulated site on the OPN promoter, using luciferase reporter assays of the promoter deletions, identified a novel NF-kappaB site (OPN/NF-kappaB). Electrophoretic mobility shift assays and chromatin immunoprecipitations (ChIP) confirmed this site to be an NF-kappaB-binding site. We also show a role of HDAC3 in suppression of OPN via OPN/NF-kappaB.
Conclusion: Our results show that BRMS1 regulates OPN transcription by abrogating NF-kappaB activation. Thus, we identify OPN, a tumor-metastasis activator, as a crucial downstream target of BRMS1. Suppression of OPN may be one of the possible underlying mechanisms of BRMS1-dependent suppression of tumor metastasis.
Figures
References
-
- Singhal H, Bautista DS, Tonkin KS, O'Malley FP, Tuck AB, Chambers AF, Harris JF. Elevated plasma osteopontin in metastatic breast cancer associated with increased tumor burden and decreased survival. Clin Cancer Res. 1997;3:605–611. - PubMed
-
- Rudland PS, Platt-Higgins A, El-Tanani M, De Silva Rudland S, Barraclough R, Winstanley JH, Howitt R, West CR. Prognostic significance of the metastasis-associated protein osteopontin in human breast cancer. Cancer Res. 2002;62:3417–3427. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
