

All four CatSper ion channel proteins are required for male fertility and sperm cell hyperactivated motility
- PMID: 17227845
- PMCID: PMC1770895
- DOI: 10.1073/pnas.0610286104
All four CatSper ion channel proteins are required for male fertility and sperm cell hyperactivated motility
Abstract
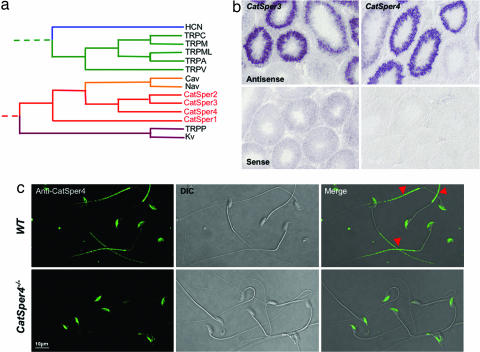
Mammalian spermatozoa become motile at ejaculation, but before they can fertilize the egg, they must acquire more thrust to penetrate the cumulus and zona pellucida. The forceful asymmetric motion of hyperactivated spermatozoa requires Ca2+ entry into the sperm tail by an alkalinization-activated voltage-sensitive Ca2+-selective current (ICatSper). Hyperactivation requires CatSper1 and CatSper2 putative ion channel genes, but the function of two other related genes (CatSper3 and CatSper4) is not known. Here we show that targeted disruption of murine CatSper3 or CatSper4 also abrogated ICatSper, sperm cell hyperactivated motility and male fertility but did not affect spermatogenesis or initial motility. Direct protein interactions among CatSpers, the sperm specificity of these proteins, and loss of ICatSper in each of the four CatSper-/- mice indicate that CatSpers are highly specialized flagellar proteins.
Conflict of interest statement
Conflict of interest: M.M.M. and J.A.C. are employees of Hydra Biosciences, a company with pending patents related to CatSper protein function. As employees, both have stock options in Hydra Biosciences. D.E.C. also owns stock in Hydra Biosciences. All other authors have no conflict of interest.
Figures



Comment in
-
Wrath of the wraiths of CatSper3 and CatSper4.Proc Natl Acad Sci U S A. 2007 Jan 23;104(4):1107-8. doi: 10.1073/pnas.0610909104. Epub 2007 Jan 16. Proc Natl Acad Sci U S A. 2007. PMID: 17227862 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
