

Palmitoylation of the TPbeta isoform of the human thromboxane A2 receptor. Modulation of G protein: effector coupling and modes of receptor internalization
- PMID: 17229546
- PMCID: PMC2680975
- DOI: 10.1016/j.cellsig.2006.12.001
Palmitoylation of the TPbeta isoform of the human thromboxane A2 receptor. Modulation of G protein: effector coupling and modes of receptor internalization
Erratum in
- Cell Signal. 2016 Mar;28(3):157
-
Corrigendum to "Palmitoylation of the TPβ isoform of the human thromboxane A2 receptor. Modulation of G protein: Effector coupling and modes of receptor internalization." [Cell Signal. 19(5) (2007) 1056-1070].Cell Signal. 2016 Mar;28(3):157. doi: 10.1016/j.cellsig.2015.11.015. Epub 2015 Dec 29. Cell Signal. 2016. PMID: 28800815 Free PMC article. No abstract available.
Abstract
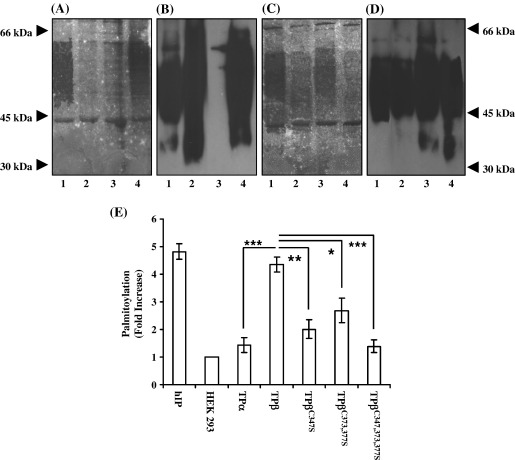
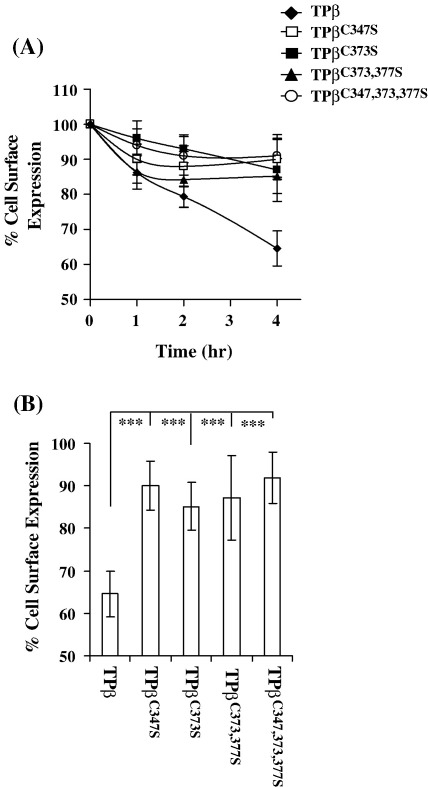
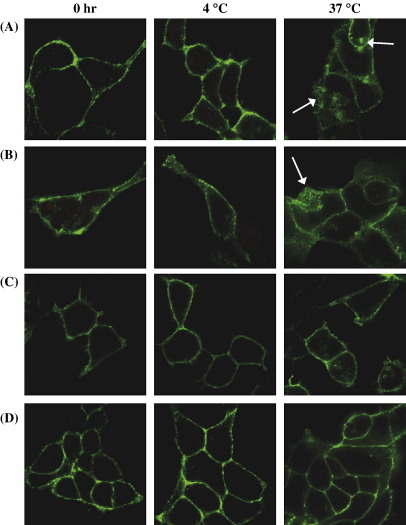
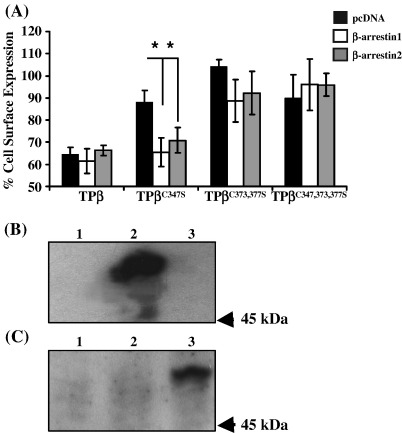
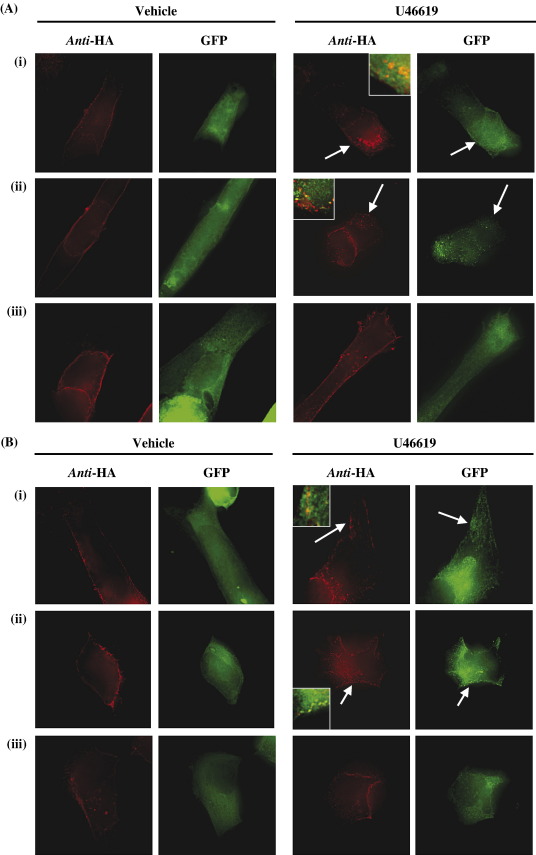
Palmitoylation is a prevalent feature amongst G protein-coupled receptors. In this study we sought to establish whether the TPalpha and TPbeta isoforms of the human prostanoid thromboxane (TX) A2 receptor (TP) are palmitoylated and to assess the functional consequences thereof. Consistent with the presence of three cysteines within its unique carboxyl-terminal domain, metabolic labelling and site-directed mutagenesis confirmed that TPbeta is palmitoylated at Cys347 and, to a lesser extent, at Cys373,377 whereas TPalpha is not palmitoylated. Impairment of palmitoylation did not affect TPbeta expression or its ligand affinity. Conversely, agonist-induced [Ca2+]i mobilization by TPbetaC347S and the non-palmitoylated TPbetaC347,373,377S, but not by TPbetaC373S or TPbetaC373,377S, was significantly reduced relative to the wild type TPbeta suggesting that palmitoylation at Cys347 is specifically required for efficient Gq/phospholipase Cbeta effector coupling. Furthermore, palmitoylation at Cys373,377 is critical for TPbeta internalization with TPbetaC373S, TPbetaC373,377S and TPbetaC347,373,377S failing to undergo either agonist-induced or temperature-dependent tonic internalization. On the other hand, whilst TPbetaC347S underwent reduced agonist-induced internalization, it underwent tonic internalization to a similar extent as TPbeta. The deficiency in agonist-induced internalization by TPbetaC347S, but not by TPbetaC373,377 nor TPbeta(C347,373,377S), was overcome by over-expression of either beta-arrestin1 or beta-arrestin2. Taken together, data herein suggest that whilst palmitoylation of TPbeta at Cys373,377 is critical for both agonist- and tonic-induced internalization, palmitoylation at Cys347 has a role in determining which pathway is followed, be it by the beta-arrestin-dependent agonist-induced pathway or by the beta-arrestin-independent tonic internalization pathway.
Figures




Similar articles
-
Homologous desensitization of signalling by the beta (beta) isoform of the human thromboxane A2 receptor.Biochim Biophys Acta. 2006 Sep;1761(9):1114-31. doi: 10.1016/j.bbalip.2006.07.012. Epub 2006 Aug 3. Biochim Biophys Acta. 2006. PMID: 16956790
-
Investigation of the role of the carboxyl-terminal tails of the alpha and beta isoforms of the human thromboxane A(2) receptor (TP) in mediating receptor:effector coupling.Biochim Biophys Acta. 2000 Apr 17;1496(2-3):164-82. doi: 10.1016/s0167-4889(00)00031-8. Biochim Biophys Acta. 2000. PMID: 10771086
-
Involvement of actin in agonist-induced endocytosis of the G protein-coupled receptor for thromboxane A2: overcoming of actin disruption by arrestin-3 but not arrestin-2.J Biol Chem. 2005 Jun 17;280(24):23215-24. doi: 10.1074/jbc.M414071200. Epub 2005 Apr 20. J Biol Chem. 2005. PMID: 15845539
-
Thromboxane A2: physiology/pathophysiology, cellular signal transduction and pharmacology.Pharmacol Ther. 2008 Apr;118(1):18-35. doi: 10.1016/j.pharmthera.2008.01.001. Epub 2008 Jan 26. Pharmacol Ther. 2008. PMID: 18374420 Review.
-
Preparing to strike: Acute events in signaling by the serpentine receptor for thromboxane A2.Pharmacol Ther. 2023 Aug;248:108478. doi: 10.1016/j.pharmthera.2023.108478. Epub 2023 Jun 13. Pharmacol Ther. 2023. PMID: 37321373 Review.
Cited by
-
The Role and Regulation of Thromboxane A2 Signaling in Cancer-Trojan Horses and Misdirection.Molecules. 2022 Sep 22;27(19):6234. doi: 10.3390/molecules27196234. Molecules. 2022. PMID: 36234768 Free PMC article. Review.
-
Differential regulation of two palmitoylation sites in the cytoplasmic tail of the beta1-adrenergic receptor.J Biol Chem. 2011 May 27;286(21):19014-23. doi: 10.1074/jbc.M110.189977. Epub 2011 Apr 4. J Biol Chem. 2011. PMID: 21464135 Free PMC article.
-
Molecular dynamics simulations reveal specific interactions of post-translational palmitoyl modifications with rhodopsin in membranes.J Am Chem Soc. 2012 Mar 7;134(9):4324-31. doi: 10.1021/ja2108382. Epub 2012 Feb 22. J Am Chem Soc. 2012. PMID: 22280374 Free PMC article.
-
Interaction of the human prostacyclin receptor with Rab11: characterization of a novel Rab11 binding domain within alpha-helix 8 that is regulated by palmitoylation.J Biol Chem. 2010 Jun 11;285(24):18709-26. doi: 10.1074/jbc.M110.106476. Epub 2010 Apr 15. J Biol Chem. 2010. Retraction in: J Biol Chem. 2018 Aug 3;293(31):12287. doi: 10.1074/jbc.W118.004807. PMID: 20395296 Free PMC article. Retracted.
-
Effects of Post-translational Modifications on Membrane Localization and Signaling of Prostanoid GPCR-G Protein Complexes and the Role of Hypoxia.J Membr Biol. 2019 Oct;252(4-5):509-526. doi: 10.1007/s00232-019-00091-4. Epub 2019 Sep 4. J Membr Biol. 2019. PMID: 31485700 Review.
References
-
- Milligan G., Parenti M., Magee A.I. Trends Biochem. Sci. 1995;20(5):181. - PubMed
-
- Dunphy J.T., Linder M.E. Biochim. Biophys. Acta. 1998;1436(1–2):245. - PubMed
-
- Bijlmakers M.J., Marsh M. Trends Cell Biol. 2003;13(1):32. - PubMed
-
- Qanbar R., Bouvier M. Pharmacol. Ther. 2003;97(1):1. - PubMed
-
- Torrecilla I., Tobin A.B. Curr. Pharm. Des. 2006;12(14):1797. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
