

A proposal for robust temperature compensation of circadian rhythms
- PMID: 17229851
- PMCID: PMC1773060
- DOI: 10.1073/pnas.0601378104
A proposal for robust temperature compensation of circadian rhythms
Abstract
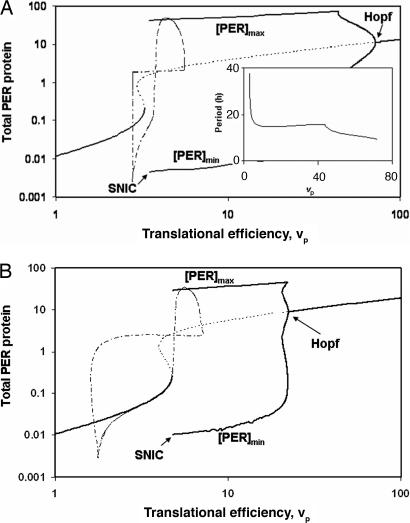
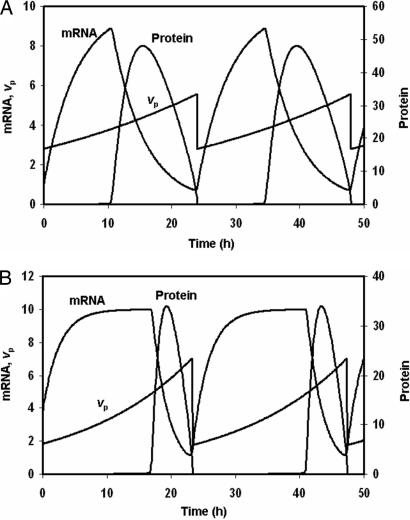
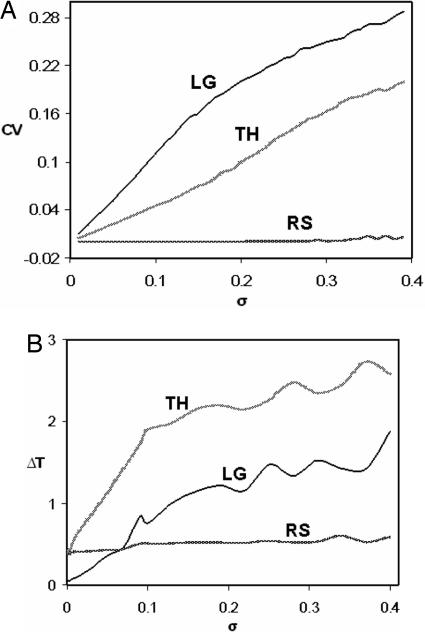
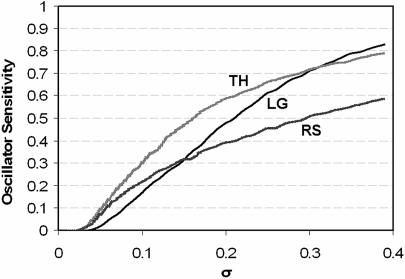
The internal circadian rhythms of cells and organisms coordinate their physiological properties to the prevailing 24-h cycle of light and dark on earth. The mechanisms generating circadian rhythms have four defining characteristics: they oscillate endogenously with period close to 24 h, entrain to external signals, suffer phase shifts by aberrant pulses of light or temperature, and compensate for changes in temperature over a range of 10 degrees C or more. Most theoretical descriptions of circadian rhythms propose that the underlying mechanism generates a stable limit cycle oscillation (in constant darkness or dim light), because limit cycles quite naturally possess the first three defining properties of circadian rhythms. On the other hand, the period of a limit cycle oscillator is typically very sensitive to kinetic rate constants, which increase markedly with temperature. Temperature compensation is therefore not a general property of limit cycle oscillations but must be imposed by some delicate balance of temperature dependent effects. However, "delicate balances" are unlikely to be robust to mutations. On the other hand, if circadian rhythms arise from a mechanism that concentrates sensitivity into a few rate constants, then the "balancing act" is likely to be more robust and evolvable. We propose a switch-like mechanism for circadian rhythms that concentrates period sensitivity in just two parameters, by forcing the system to alternate between a stable steady state and a stable limit cycle.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Temperature compensation and entrainment in circadian rhythms.Phys Biol. 2012 Jun;9(3):036011. doi: 10.1088/1478-3975/9/3/036011. Epub 2012 Jun 8. Phys Biol. 2012. PMID: 22683844
-
Calculating activation energies for temperature compensation in circadian rhythms.Phys Biol. 2011 Oct;8(5):056007. doi: 10.1088/1478-3975/8/5/056007. Epub 2011 Sep 2. Phys Biol. 2011. PMID: 21891835
-
Thermocyclic and photocyclic entrainment of circadian locomotor activity rhythms in sleepy lizards, Tiliqua rugosa.Chronobiol Int. 2009 Oct;26(7):1369-88. doi: 10.3109/07420520903412392. Chronobiol Int. 2009. PMID: 19916837
-
The effects of social defeat and other stressors on the expression of circadian rhythms.Stress. 2002 Feb;5(1):15-22. doi: 10.1080/102538902900012323. Stress. 2002. PMID: 12171763 Review.
-
Circadian rhythms: influences of internal and external factors on the period measured in constant conditions.Z Tierpsychol. 1979 Mar;49(3):225-49. doi: 10.1111/j.1439-0310.1979.tb00290.x. Z Tierpsychol. 1979. PMID: 386643 Review.
Cited by
-
Molecular characterization of the transition to mid-life in Caenorhabditis elegans.Age (Dordr). 2013 Jun;35(3):689-703. doi: 10.1007/s11357-012-9401-2. Epub 2012 May 20. Age (Dordr). 2013. PMID: 22610697 Free PMC article.
-
Non-sinusoidal Waveform in Temperature-Compensated Circadian Oscillations.Biophys J. 2019 Feb 19;116(4):741-751. doi: 10.1016/j.bpj.2018.12.022. Epub 2019 Jan 15. Biophys J. 2019. PMID: 30712786 Free PMC article.
-
Biological switches and clocks.J R Soc Interface. 2008 Aug 6;5 Suppl 1(Suppl 1):S1-8. doi: 10.1098/rsif.2008.0179.focus. J R Soc Interface. 2008. PMID: 18522926 Free PMC article.
-
Multi-scale modeling of the circadian modulation of learning and memory.PLoS One. 2019 Jul 19;14(7):e0219915. doi: 10.1371/journal.pone.0219915. eCollection 2019. PLoS One. 2019. PMID: 31323054 Free PMC article.
-
Systematic analysis of negative and positive feedback loops for robustness and temperature compensation in circadian rhythms.NPJ Syst Biol Appl. 2023 Feb 11;9(1):5. doi: 10.1038/s41540-023-00268-7. NPJ Syst Biol Appl. 2023. PMID: 36774353 Free PMC article.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
