

Telomerase abrogation dramatically accelerates TRF2-induced epithelial carcinogenesis
- PMID: 17234886
- PMCID: PMC1770903
- DOI: 10.1101/gad.406207
Telomerase abrogation dramatically accelerates TRF2-induced epithelial carcinogenesis
Abstract
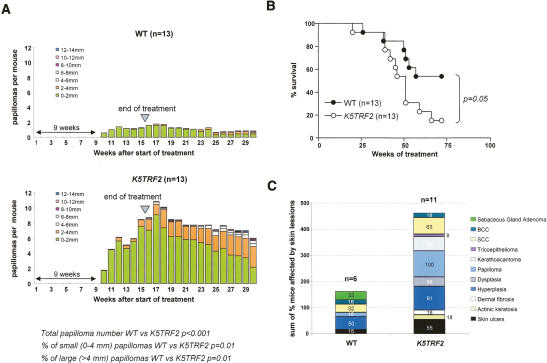
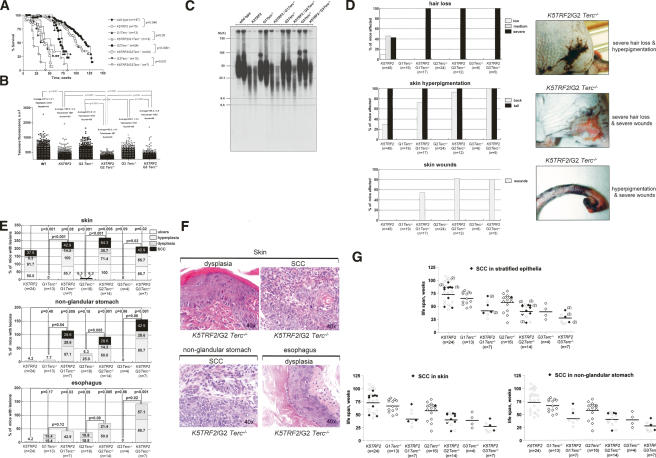
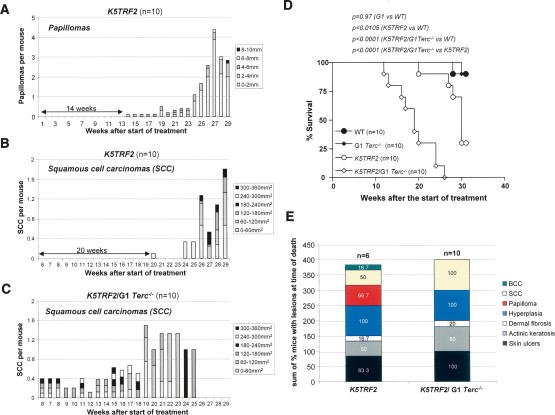
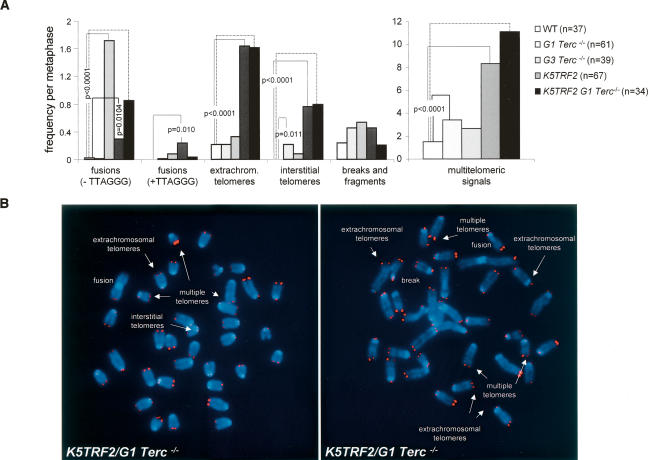
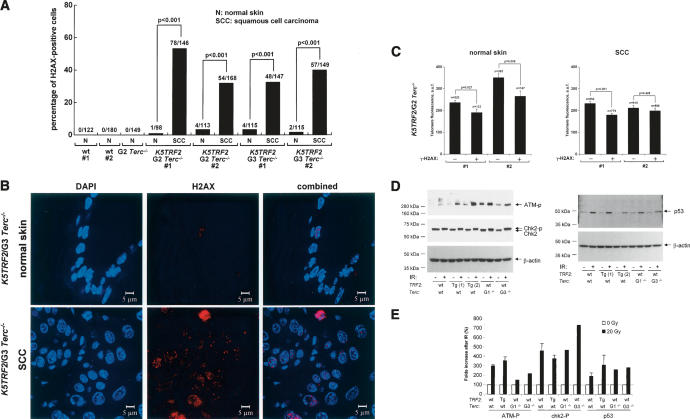
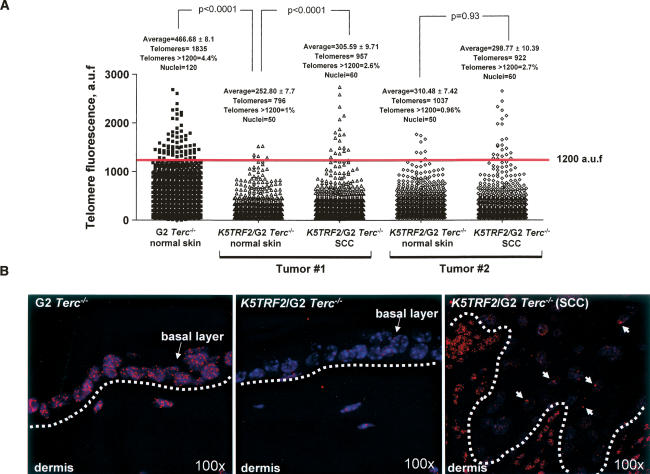
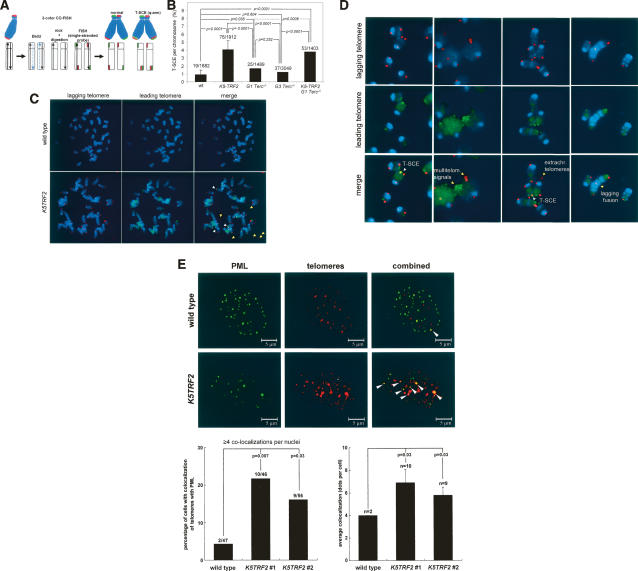
TRF2 is a telomere-binding protein with roles in telomere protection and telomere-length regulation. The fact that TRF2 is up-regulated in some human tumors suggests a role of TRF2 in cancer. Mice that overexpress TRF2 in the skin, K5TRF2 mice, show critically short telomeres and are susceptible to UV-induced carcinogenesis as a result of deregulated XPF/ERCC1 activity, a nuclease involved in UV damage repair. Here we demonstrate that, when in combination with telomerase deficiency, TRF2 acts as a very potent oncogene in vivo. In particular, we show that telomerase deficiency dramatically accelerates TRF2-induced epithelial carcinogenesis in K5TRF2/Terc-/- mice, coinciding with increased chromosomal instability and DNA damage. Telomere recombination is also increased in these mice, suggesting that TRF2 favors the activation of alternative telomere maintenance mechanisms. Together, these results demonstrate that TRF2 increased expression is a potent oncogenic event that along with telomerase deficiency accelerates carcinogenesis, coincidental with a derepression of telomere recombination. These results are of particular relevance given that TRF2 is up-regulated in some human cancers. Furthermore, these data suggest that telomerase inhibition might not be effective to cease the growth of TRF2-overexpressing tumors.
Figures
References
-
- Artandi S.E., Chang S., Lee S.L., Alson S., Gottlieb G.J., Chin L., DePinho R.A., Chang S., Lee S.L., Alson S., Gottlieb G.J., Chin L., DePinho R.A., Lee S.L., Alson S., Gottlieb G.J., Chin L., DePinho R.A., Alson S., Gottlieb G.J., Chin L., DePinho R.A., Gottlieb G.J., Chin L., DePinho R.A., Chin L., DePinho R.A., DePinho R.A. Telomere dysfunction promotes non-reciprocal translocations and epithelial cancers in mice. Nature. 2000;406:641–645. - PubMed
-
- Balmain A., Ramsden M., Bowden G.T., Smith J., Ramsden M., Bowden G.T., Smith J., Bowden G.T., Smith J., Smith J. Activation of the mouse Harvey-ras gene in chemically induced benign skin papillomas. Nature. 1984;307:658–660. - PubMed
-
- Bechter O.E., Zou Y., Walker W., Wright W.E., Shay J.W., Zou Y., Walker W., Wright W.E., Shay J.W., Walker W., Wright W.E., Shay J.W., Wright W.E., Shay J.W., Shay J.W. Telomeric recombination in mismatch repair deficient human colon cancer cells and telomerase inhibition. Cancer Res. 2004;64:3444–3451. - PubMed
-
- Biroccio A., Rizzo A., Elli R., Koering C.E., Belleville A., Benassi B., Leonetti C., Stevens M.F., D’Incalci M., Zupi G., Rizzo A., Elli R., Koering C.E., Belleville A., Benassi B., Leonetti C., Stevens M.F., D’Incalci M., Zupi G., Elli R., Koering C.E., Belleville A., Benassi B., Leonetti C., Stevens M.F., D’Incalci M., Zupi G., Koering C.E., Belleville A., Benassi B., Leonetti C., Stevens M.F., D’Incalci M., Zupi G., Belleville A., Benassi B., Leonetti C., Stevens M.F., D’Incalci M., Zupi G., Benassi B., Leonetti C., Stevens M.F., D’Incalci M., Zupi G., Leonetti C., Stevens M.F., D’Incalci M., Zupi G., Stevens M.F., D’Incalci M., Zupi G., D’Incalci M., Zupi G., Zupi G., et al. TRF2 inhibition triggers apoptosis and reduces tumorigenicity of human melanoma cells. Eur. J. Cancer. 2006;42:1881–1888. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous