

Carbon cycling in Anabaena sp. PCC 7120. Sucrose synthesis in the heterocysts and possible role in nitrogen fixation
- PMID: 17237189
- PMCID: PMC1820908
- DOI: 10.1104/pp.106.091736
Carbon cycling in Anabaena sp. PCC 7120. Sucrose synthesis in the heterocysts and possible role in nitrogen fixation
Abstract
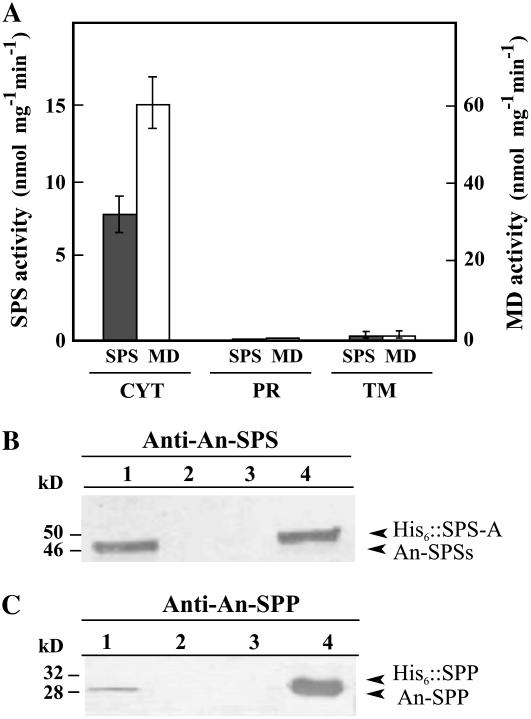
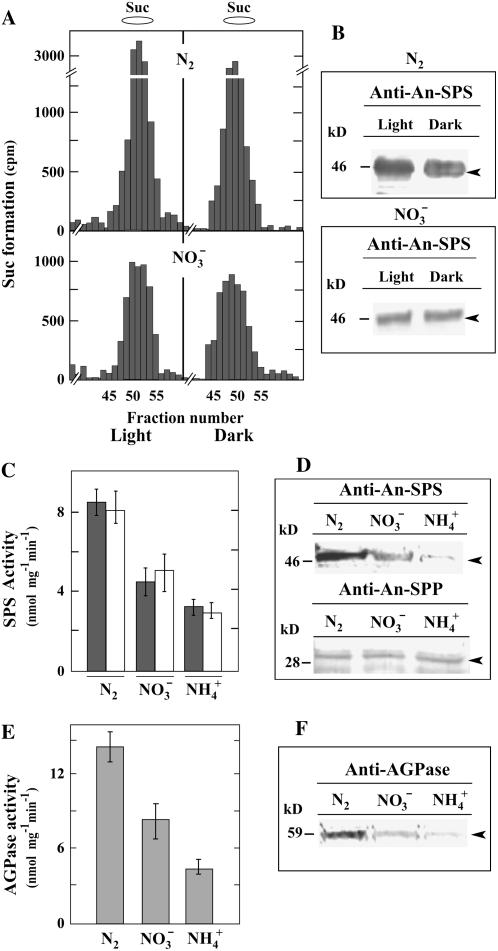
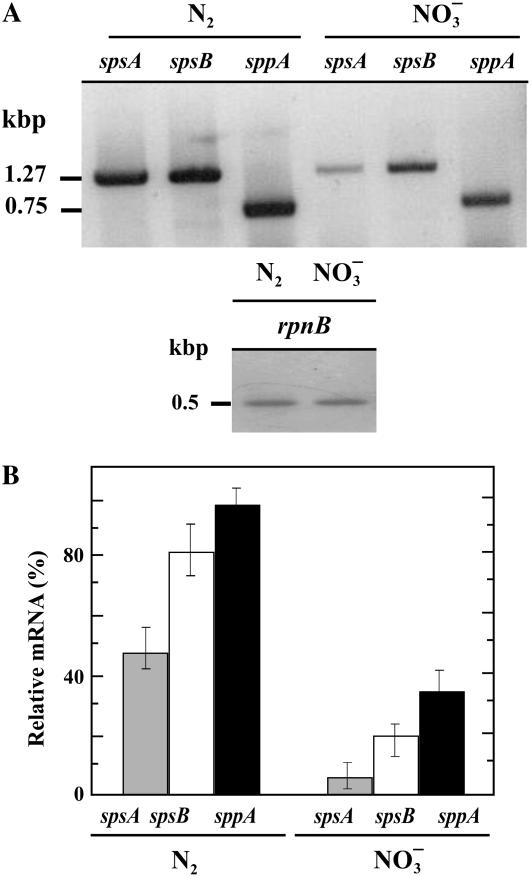
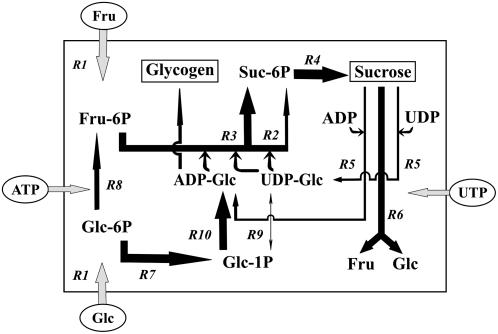
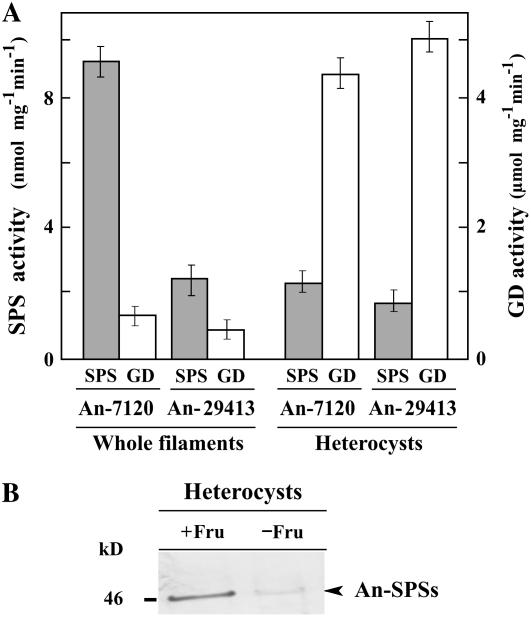
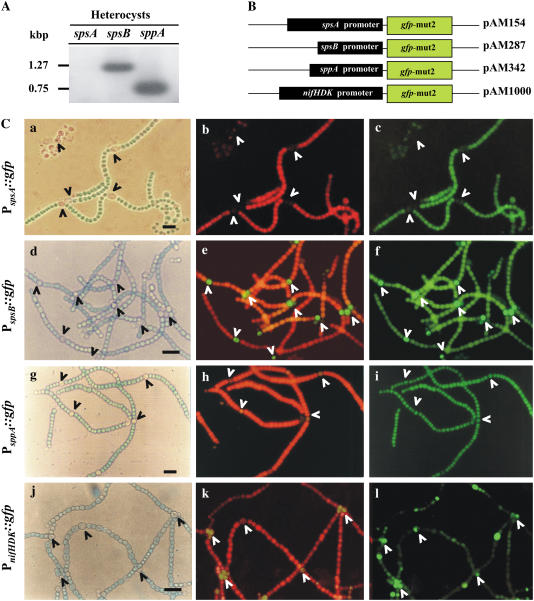
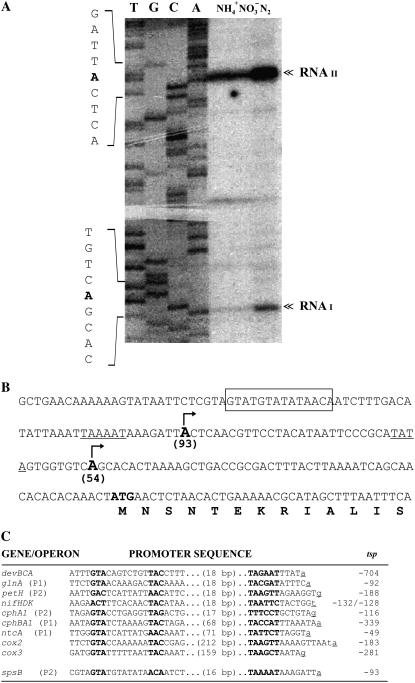
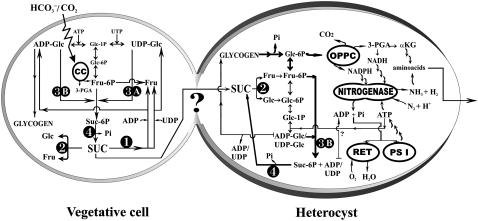
Nitrogen (N) available to plants mostly originates from N(2) fixation carried out by prokaryotes. Certain cyanobacterial species contribute to this energetically expensive process related to carbon (C) metabolism. Several filamentous strains differentiate heterocysts, specialized N(2)-fixing cells. To understand how C and N metabolism are regulated in photodiazotrophically grown organisms, we investigated the role of sucrose (Suc) biosynthesis in N(2) fixation in Anabaena sp. PCC 7120 (also known as Nostoc sp. PCC 7120). The presence of two Suc-phosphate synthases (SPS), SPS-A and SPS-B, directly involved in Suc synthesis with different glucosyl donor specificity, seems to be important in the N(2)-fixing filament. Measurement of enzyme activity and polypeptide levels plus reverse transcription-polymerase chain reaction experiments showed that total SPS expression is greater in cells grown in N(2) versus combined N conditions. Only SPS-B, however, was seen to be active in the heterocyst, as confirmed by analysis of green fluorescent protein reporters. SPS-B gene expression is likely controlled at the transcriptional initiation level, probably in relation to a global N regulator. Metabolic control analysis indicated that the metabolism of glycogen and Suc is likely interconnected in N(2)-fixing filaments. These findings suggest that N(2) fixation may be spatially compatible with Suc synthesis and support the role of the disaccharide as an intermediate in the reduced C flux in heterocyst-forming cyanobacteria.
Figures
References
-
- Barreiro R (1999) Metabolic control analysis of carbon pathways. PhD thesis. UMI Dissertation Services, Ann Arbor, MI
-
- Berman-Frank I, Lundgren P, Falkowski P (2003) Nitrogen fixation and photosynthetic oxygen evolution in cyanobacteria. Res Microbiol 154 157–164 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
