

Stiffness and fraction of Myosin motors responsible for active force in permeabilized muscle fibers from rabbit psoas
- PMID: 17237201
- PMCID: PMC1864836
- DOI: 10.1529/biophysj.106.099549
Stiffness and fraction of Myosin motors responsible for active force in permeabilized muscle fibers from rabbit psoas
Abstract
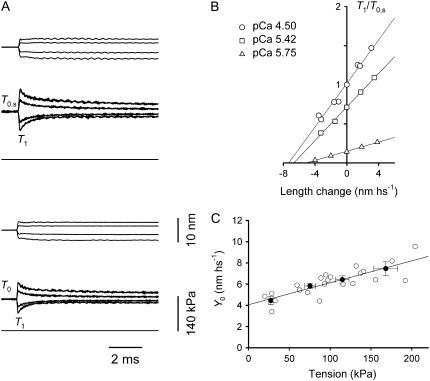
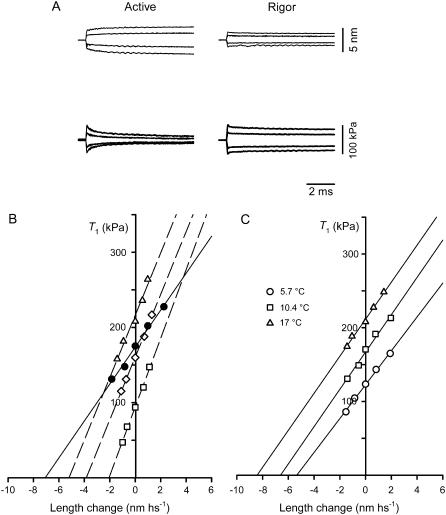
The stiffness of the single myosin motor (epsilon) is determined in skinned fibers from rabbit psoas muscle by both mechanical and thermodynamic approaches. Changes in the elastic strain of the half-sarcomere (hs) are measured by fast mechanics both in rigor, when all myosin heads are attached, and during active contraction, with the isometric force (T0) modulated by changing either [Ca2+] or temperature. The hs compliance is 43.0+/-0.8 nm MPa-1 in isometric contraction at saturating [Ca2+], whereas in rigor it is 28.2+/-1.1 nm MPa-1. The equivalent compliance of myofilaments is 21.0+/-3.3 nm MPa-1. Accordingly, the stiffness of the ensemble of myosin heads attached in the hs is 45.5+/-1.7 kPa nm-1 in isometric contraction at saturating [Ca2+] (e0), and in rigor (er) it rises to 138.9+/-21.2 kPa nm-1. Epsilon, calculated from er and the lattice molecular dimensions, is 1.21+/-0.18 pN nm-1. epsilon estimated, using a thermodynamic approach, from the relation of T0 at saturating [Ca2+] versus the reciprocal of absolute temperature is 1.25+/-0.14 pN nm-1, similar to that estimated for fibers in rigor. Consequently, the ratio e0/er (0.33+/-0.05) can be used to estimate the fraction of attached heads during isometric contraction at saturating [Ca2+]. If the osmotic agent dextran T-500 (4 g/100 ml) is used to reduce the lateral filament spacing of the relaxed fiber to the value before skinning, both e0 and er increase by approximately 40%. Epsilon becomes approximately 1.7 pN nm-1 and the fraction and the force of myosin heads attached in the isometric contraction remain the same as before dextran application. The finding that the fraction of myosin heads attached to actin in an isometric contraction is 0.33 rules out the hypothesis of multiple mechanical cycles per ATP hydrolyzed.
Figures
References
-
- Howard, J. 2001. Mechanics of Motor Protein and the Cytoskeleton. Sinauer, Sunderland, MA.
-
- Huxley, A. F. 1957. Muscle structure and theories of contraction. Prog. Biophys. Biophys. Chem. 7:255–318. - PubMed
-
- Huxley, A. F., and R. M. Simmons. 1971. Proposed mechanism of force generation in striated muscle. Nature. 233:533–538. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
