

S locus genes and the evolution of self-fertility in Arabidopsis thaliana
- PMID: 17237349
- PMCID: PMC1820967
- DOI: 10.1105/tpc.106.048199
S locus genes and the evolution of self-fertility in Arabidopsis thaliana
Abstract
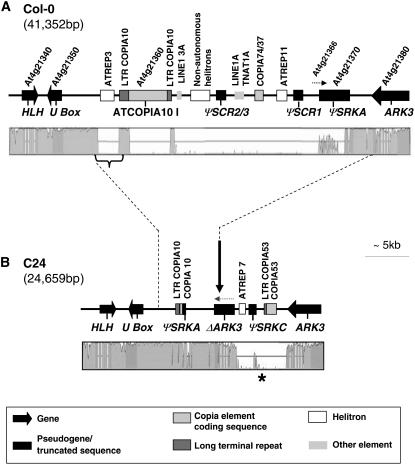
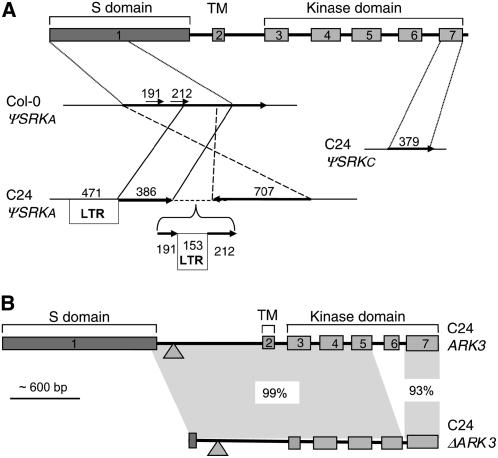
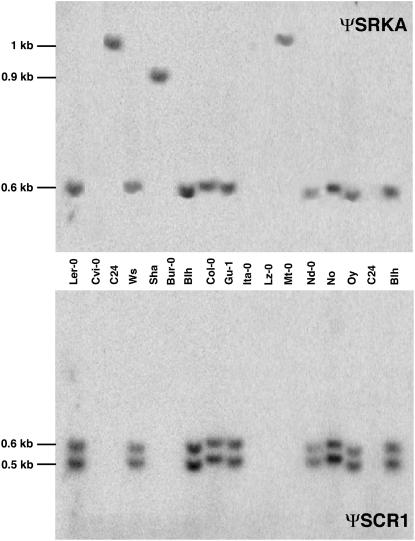
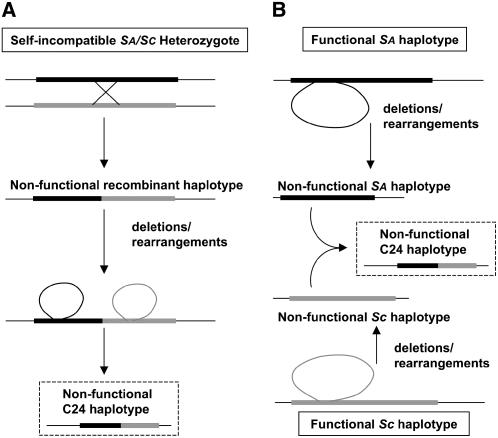
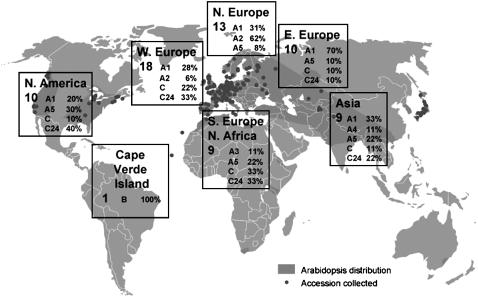
Loss of self-incompatibility (SI) in Arabidopsis thaliana was accompanied by inactivation of genes required for SI, including S-LOCUS RECEPTOR KINASE (SRK) and S-LOCUS CYSTEINE-RICH PROTEIN (SCR), coadapted genes that constitute the SI specificity-determining S haplotype. Arabidopsis accessions are polymorphic for PsiSRK and PsiSCR, but it is unknown if the species harbors structurally different S haplotypes, either representing relics of ancestral functional and structurally heteromorphic S haplotypes or resulting from decay concomitant with or subsequent to the switch to self-fertility. We cloned and sequenced the S haplotype from C24, in which self-fertility is due solely to S locus inactivation, and show that this haplotype was produced by interhaplotypic recombination. The highly divergent organization and sequence of the C24 and Columbia-0 (Col-0) S haplotypes demonstrate that the A. thaliana S locus underwent extensive structural remodeling in conjunction with a relaxation of selective pressures that once preserved the integrity and linkage of coadapted SRK and SCR alleles. Additional evidence for this process was obtained by assaying 70 accessions for the presence of C24- or Col-0-specific sequences. Furthermore, analysis of SRK and SCR polymorphisms in these accessions argues against the occurrence of a selective sweep of a particular allele of SCR, as previously proposed.
Figures
References
-
- Bakker, E.G., Stahl, E.A., Toomajian, C., Nordborg, M., and Kreitman, M. (2006). Distribution of genetic variation within and among local populations of Arabidopsis thaliana over its species range. Mol. Ecol. 15 1405–1418. - PubMed
-
- Bechsgaard, J., Castric, V., Charlesworth, D., Vekemans, X., and Schierup, M.H. (2006). The transition to self-compatibility in Arabidopsis thaliana and evolution within S-haplotypes over 10 million years. Mol. Biol. Evol. 23 1741–1750. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
