

Interactions among PIN-FORMED and P-glycoprotein auxin transporters in Arabidopsis
- PMID: 17237354
- PMCID: PMC1820964
- DOI: 10.1105/tpc.106.040782
Interactions among PIN-FORMED and P-glycoprotein auxin transporters in Arabidopsis
Abstract
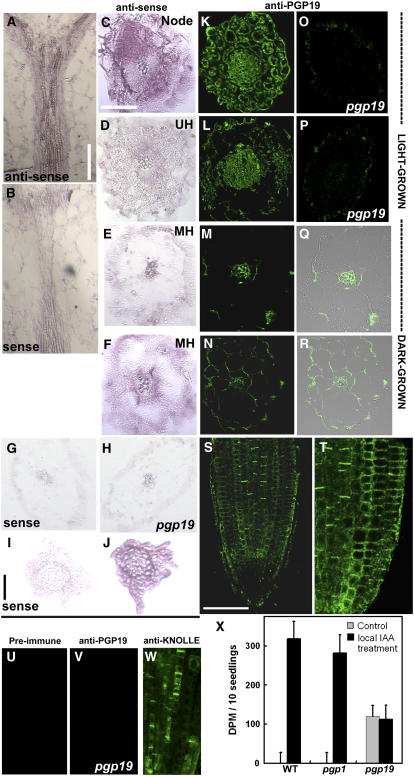
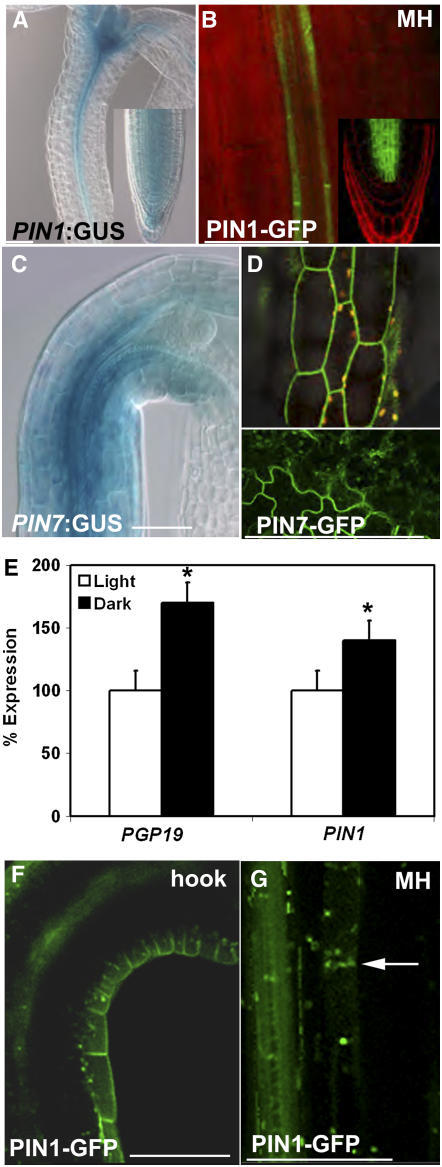
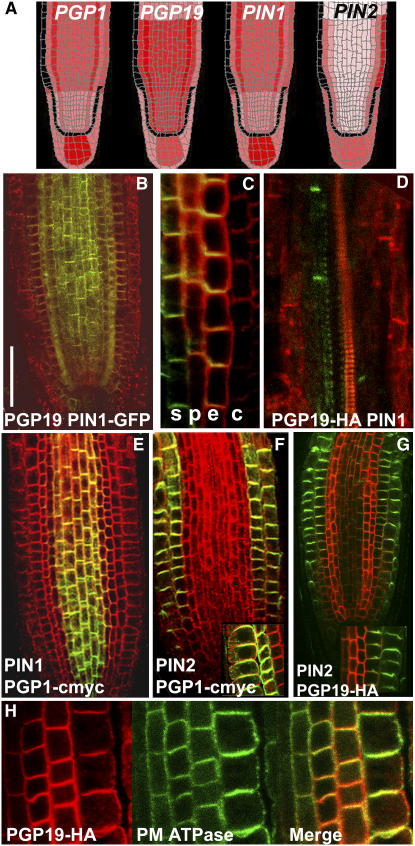
Directional transport of the phytohormone auxin is established primarily at the point of cellular efflux and is required for the establishment and maintenance of plant polarity. Studies in whole plants and heterologous systems indicate that PIN-FORMED (PIN) and P-glycoprotein (PGP) transport proteins mediate the cellular efflux of natural and synthetic auxins. However, aromatic anion transport resulting from PGP and PIN expression in nonplant systems was also found to lack the high level of substrate specificity seen in planta. Furthermore, previous reports that PGP19 stabilizes PIN1 on the plasma membrane suggested that PIN-PGP interactions might regulate polar auxin efflux. Here, we show that PGP1 and PGP19 colocalized with PIN1 in the shoot apex in Arabidopsis thaliana and with PIN1 and PIN2 in root tissues. Specific PGP-PIN interactions were seen in yeast two-hybrid and coimmunoprecipitation assays. PIN-PGP interactions appeared to enhance transport activity and, to a greater extent, substrate/inhibitor specificities when coexpressed in heterologous systems. By contrast, no interactions between PGPs and the AUXIN1 influx carrier were observed. Phenotypes of pin and pgp mutants suggest discrete functional roles in auxin transport, but pin pgp mutants exhibited phenotypes that are both additive and synergistic. These results suggest that PINs and PGPs characterize coordinated, independent auxin transport mechanisms but also function interactively in a tissue-specific manner.
Figures




References
-
- Benkova, E., Michniewicz, M., Sauer, M., Teichmann, T., Seifertova, D., Jurgens, G., and Friml, J. (2003). Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell 115 591–602. - PubMed
-
- Bennett, M.J., Marchant, A., Green, H.G., May, S.T., Ward, S.P., Millner, P.A., Walker, A.R., Schulz, B., and Feldmann, K.A. (1996). Arabidopsis AUX1 gene: A permease-like regulator of root gravitropism. Science 273 948–950. - PubMed
-
- Birnbaum, K., Shasha, D.E., Wang, J.Y., Jung, J.W., Lambert, G.M., Galbraith, D.W., and Benfey, P.N. (2003). A gene expression map of the Arabidopsis root. Science 302 1956–1960. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
