

The role of nonhomologous end-joining components in telomere metabolism in Kluyveromyces lactis
- PMID: 17237517
- PMCID: PMC1840097
- DOI: 10.1534/genetics.106.067447
The role of nonhomologous end-joining components in telomere metabolism in Kluyveromyces lactis
Abstract
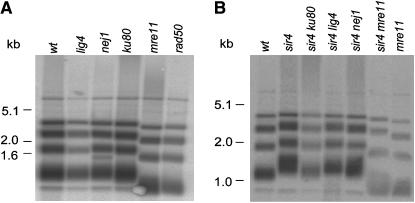
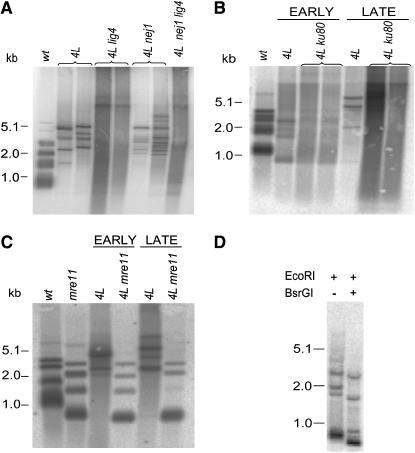
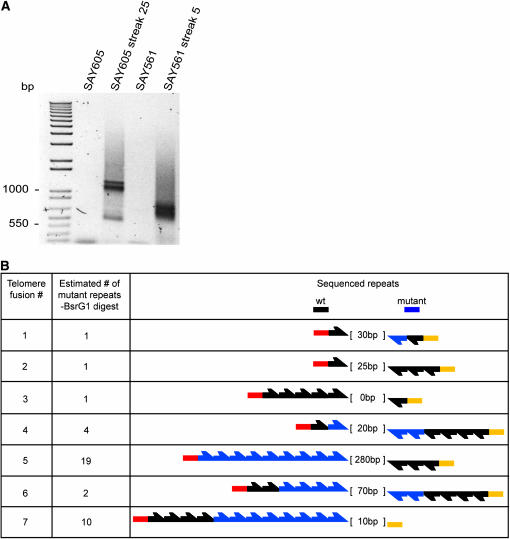
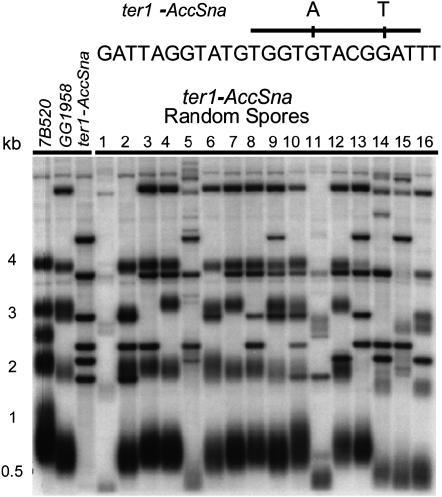
The relationship between telomeres and nonhomologous end-joining (NHEJ) is paradoxical, as NHEJ proteins are part of the telomere cap, which serves to differentiate telomeres from DNA double-strand breaks. We explored these contradictory functions for NHEJ proteins by investigating their role in Kluyveromyces lactis telomere metabolism. The ter1-4LBsr allele of the TER1 gene resulted in the introduction of sequence altered telomeric repeats and subsequent telomere-telomere fusions (T-TFs). In this background, Lig4 and Ku80 were necessary for T-TFs to form. Nej1, essential for NHEJ at internal positions, was not. Hence, T-TF formation was mediated by an unusual NHEJ mechanism. Rad50 and mre11 strains exhibited stable short telomeres, suggesting that Rad50 and Mre11 were required for telomerase recruitment. Introduction of the ter1-4LBsr allele into these strains failed to result in telomere elongation as normally observed with the ter1-4LBsr allele. Thus, the role of Rad50 and Mre11 in the formation of T-TFs was unclear. Furthermore, rad50 and mre11 mutants had highly increased subtelomeric recombination rates, while ku80 and lig4 mutants displayed moderate increases. Ku80 mutant strains also contained extended single-stranded 3' telomeric overhangs. We concluded that NHEJ proteins have multiple roles at telomeres, mediating fusions of mutant telomeres and ensuring end protection of normal telomeres.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
