

In vitro derived dendritic cells trans-infect CD4 T cells primarily with surface-bound HIV-1 virions
- PMID: 17238285
- PMCID: PMC1779297
- DOI: 10.1371/journal.ppat.0030004
In vitro derived dendritic cells trans-infect CD4 T cells primarily with surface-bound HIV-1 virions
Abstract
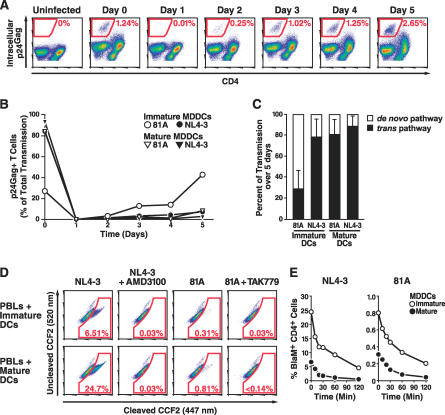
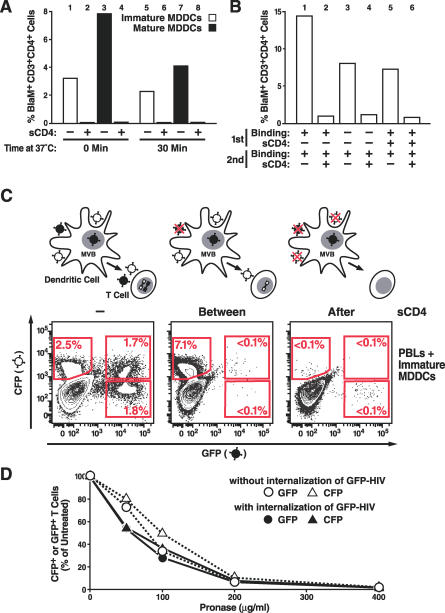
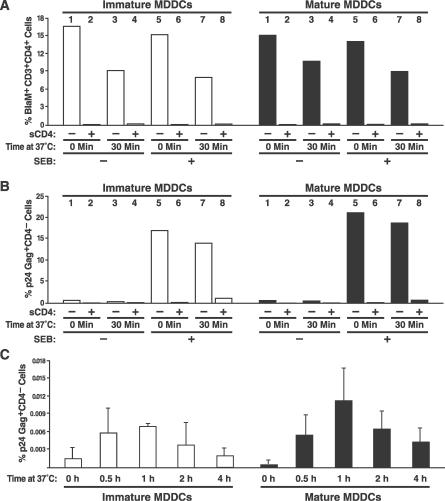
In the prevailing model of HIV-1 trans-infection, dendritic cells (DCs) capture and internalize intact virions and transfer these virions to interacting T cells at the virological synapse. Here, we show that HIV-1 virions transmitted in trans from in vitro derived DCs to T cells principally originate from the surface of DCs. Selective neutralization of surface-bound virions abrogated trans-infection by monocyte-derived DCs and CD34-derived Langerhans cells. Under conditions mimicking antigen recognition by the interacting T cells, most transferred virions still derived from the cell surface, although a few were transferred from an internal compartment. Our findings suggest that attachment inhibitors could neutralize trans-infection of T cells by DCs in vivo.
Conflict of interest statement
Figures
References
-
- Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature. 1998;392:245–252. - PubMed
-
- Cameron PU, Freudenthal PS, Barker JM, Gezelter S, Inaba K, et al. Dendritic cells exposed to human immunodeficiency virus type-1 transmit a vigorous cytopathic infection to CD4+ T cells. Science. 1992;257:383–387. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
