

SecY alterations that impair membrane protein folding and generate a membrane stress
- PMID: 17242069
- PMCID: PMC2063957
- DOI: 10.1083/jcb.200611121
SecY alterations that impair membrane protein folding and generate a membrane stress
Abstract
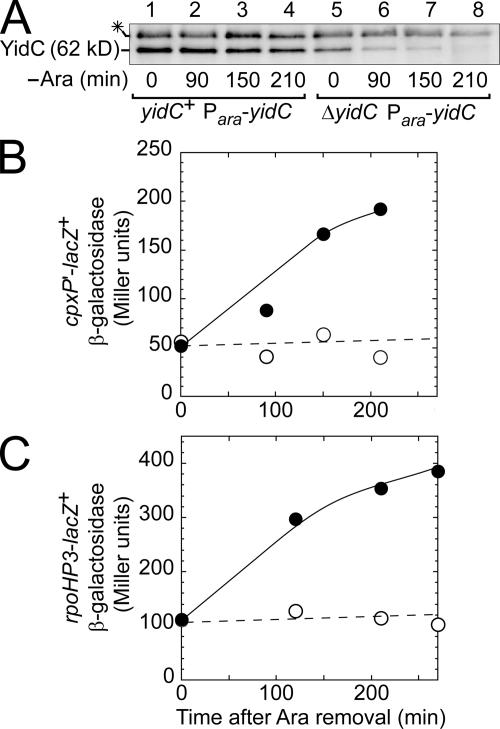
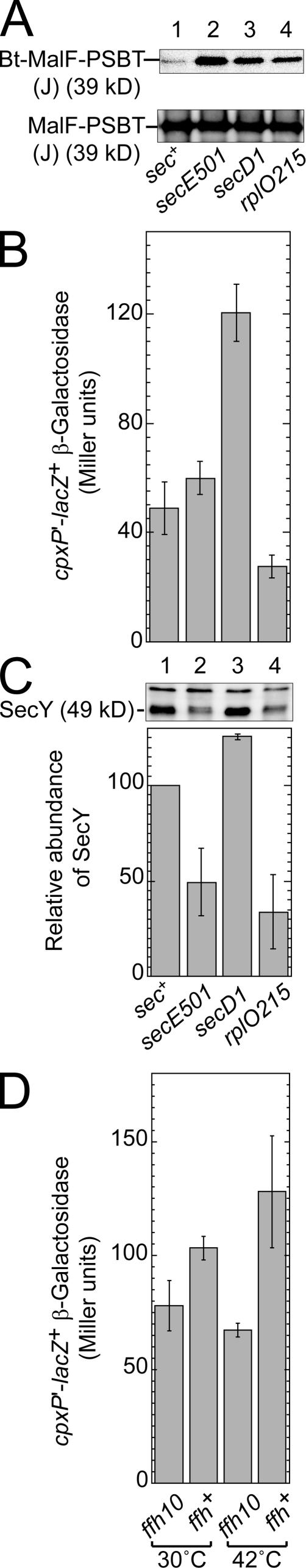
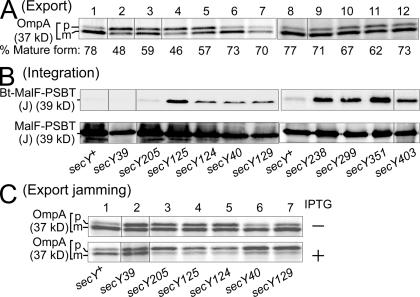
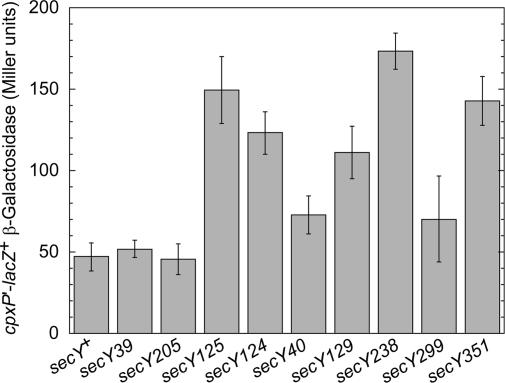
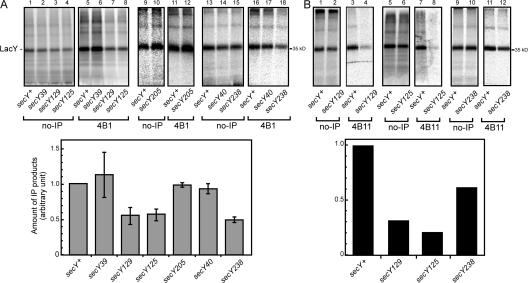
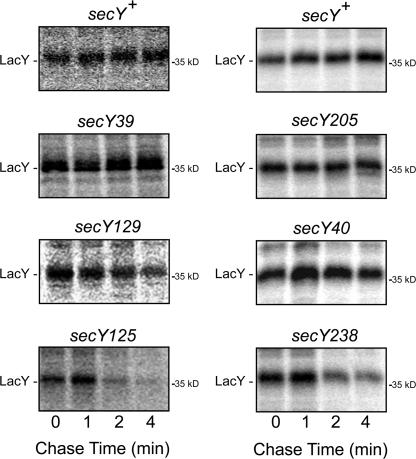
We report on a class of Escherichia coli SecY mutants that impair membrane protein folding. The mutants also up-regulate the Cpx/sigma(E) stress response pathways. Similar stress induction was also observed in response to a YidC defect in membrane protein biogenesis but not in response to the signal recognition particle-targeting defect or in response to a simple reduction in the abundance of the translocon. Together with the previous contention that the Cpx system senses a protein abnormality not only at periplasmic and outer membrane locations but also at the plasma membrane, abnormal states of membrane proteins are postulated to be generated in these secY mutants. In support of this notion, in vitro translation, membrane integration, and folding of LacY reveal that mutant membrane vesicles allow the insertion of LacY but not subsequent folding into a normal conformation recognizable by conformation-specific antibodies. The results demonstrate that normal SecY function is required for the folding of membrane proteins after their insertion into the translocon.
Figures
References
-
- Abramson, J., I. Smirnova, V. Kasho, G. Verner, H.R. Kaback, and S. Iwata. 2003. Structure and mechanism of the lactose permease of Escherichia coli. Science. 301:610–615. - PubMed
-
- Akiyama, Y., and K. Ito. 1989. Export of Escherichia coli alkaline phosphatase attached to an integral membrane protein, SecY. J. Biol. Chem. 264:437–442. - PubMed
-
- Akiyama, Y., and K. Ito. 1990. SecY protein, a membrane-embedded secretion factor of E. coli, is cleaved by the OmpT protease in vitro. Biochem. Biophys. Res. Commun. 167:711–715. - PubMed
