

A mechanosensory system controls cell shape changes during mitosis
- PMID: 17245114
- PMCID: PMC4638380
- DOI: 10.4161/cc.6.1.3674
A mechanosensory system controls cell shape changes during mitosis
Abstract
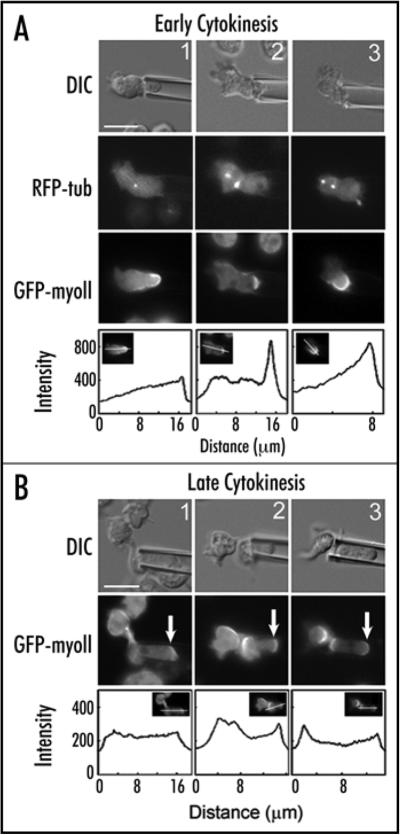
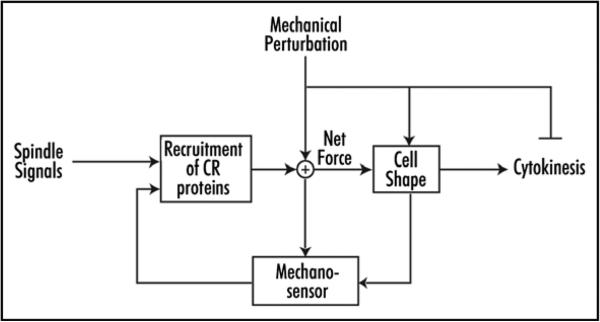
Essential life processes are heavily controlled by a variety of positive and negative feedback systems. Cytokinesis failure, ultimately leading to aneuploidy, is appreciated as an early step in tumor formation in mammals and is deleterious for all cells. Further, the growing list of cancer predisposition mutations includes a number of genes whose proteins control mitosis and/or cytokinesis. Cytokinesis shape control is also an important part of pattern formation and cell-type specialization during multi-cellular development. Inherently mechanical, we hypothesized that mechanosensing and mechanical feedback are fundamental for cytokinesis shape regulation. Using mechanical perturbation, we identified a mechanosensory control system that monitors shape progression during cytokinesis. In this review, we summarize these findings and their implications for cytokinesis regulation and for understanding the cytoskeletal system architecture that governs shape control.
Figures


Similar articles
-
Polar actomyosin contractility destabilizes the position of the cytokinetic furrow.Nature. 2011 Aug 7;476(7361):462-6. doi: 10.1038/nature10286. Nature. 2011. PMID: 21822289
-
Mitosis-specific mechanosensing and contractile-protein redistribution control cell shape.Curr Biol. 2006 Oct 10;16(19):1962-7. doi: 10.1016/j.cub.2006.08.027. Curr Biol. 2006. PMID: 17027494 Free PMC article.
-
Mechanical stress and network structure drive protein dynamics during cytokinesis.Curr Biol. 2015 Mar 2;25(5):663-70. doi: 10.1016/j.cub.2015.01.025. Epub 2015 Feb 19. Curr Biol. 2015. PMID: 25702575 Free PMC article.
-
Separation anxiety: stress, tension and cytokinesis.Exp Cell Res. 2012 Jul 15;318(12):1428-34. doi: 10.1016/j.yexcr.2012.03.028. Epub 2012 Mar 31. Exp Cell Res. 2012. PMID: 22487096 Free PMC article. Review.
-
Cytokinesis: Robust cell shape regulation.Semin Cell Dev Biol. 2016 May;53:39-44. doi: 10.1016/j.semcdb.2015.10.023. Epub 2015 Oct 19. Semin Cell Dev Biol. 2016. PMID: 26481973 Free PMC article. Review.
Cited by
-
Polar actomyosin contractility destabilizes the position of the cytokinetic furrow.Nature. 2011 Aug 7;476(7361):462-6. doi: 10.1038/nature10286. Nature. 2011. PMID: 21822289
-
The spatial and mechanical challenges of female meiosis.Mol Reprod Dev. 2011 Oct-Nov;78(10-11):769-77. doi: 10.1002/mrd.21358. Epub 2011 Jul 19. Mol Reprod Dev. 2011. PMID: 21774026 Free PMC article. Review.
-
Mitotic spindle orients perpendicular to the forces imposed by dynamic shear.PLoS One. 2011;6(12):e28965. doi: 10.1371/journal.pone.0028965. Epub 2011 Dec 29. PLoS One. 2011. PMID: 22220200 Free PMC article.
-
Punctuated actin contractions during convergent extension and their permissive regulation by the non-canonical Wnt-signaling pathway.J Cell Sci. 2011 Feb 15;124(Pt 4):635-46. doi: 10.1242/jcs.067579. Epub 2011 Jan 25. J Cell Sci. 2011. PMID: 21266466 Free PMC article.
-
Distinct apical and basolateral membrane requirements for stretch-induced membrane traffic at the apical surface of bladder umbrella cells.Mol Biol Cell. 2009 Jan;20(1):282-95. doi: 10.1091/mbc.e08-04-0439. Epub 2008 Nov 5. Mol Biol Cell. 2009. PMID: 18987341 Free PMC article.
References
-
- Fujiwara T, Bandi M, Nitta M, Ivanova EV, Bronson RT, Pellman D. Cytokinesis failure generating tetraploids promotes tumorigenesis in p53-null cells. Nature. 2005;437:1043–7. - PubMed
-
- Giet R, Petretti C, Prigent C. Aurora kinases, aneuploidy and cancer, a coincidence or a real link? Trends Cell Biol. 2005;15:241–50. - PubMed
-
- Gisselsson D. Mitotic instability in cancer: Is there a method in the madness? Cell Cycle. 2005;4:1007–10. - PubMed
-
- Wooster R, Neuhausen SL, Mangion J, Quirk Y, Ford D, Collins N, Nguyen K, Seal S, Tran T, Averill D. Localization of a breast cancer susceptibility gene, BRCA2, to chromosome 13q12-13. Science. 1994;265:2088–90. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources