

Stochastic amplification in epidemics
- PMID: 17251128
- PMCID: PMC2373404
- DOI: 10.1098/rsif.2006.0192
Stochastic amplification in epidemics
Abstract
The role of stochasticity and its interplay with nonlinearity are central current issues in studies of the complex population patterns observed in nature, including the pronounced oscillations of wildlife and infectious diseases. The dynamics of childhood diseases have provided influential case studies to develop and test mathematical models with practical application to epidemiology, but are also of general relevance to the central question of whether simple nonlinear systems can explain and predict the complex temporal and spatial patterns observed in nature outside laboratory conditions. Here, we present a stochastic theory for the major dynamical transitions in epidemics from regular to irregular cycles, which relies on the discrete nature of disease transmission and low spatial coupling. The full spectrum of stochastic fluctuations is derived analytically to show how the amplification of noise varies across these transitions. The changes in noise amplification and coherence appear robust to seasonal forcing, questioning the role of seasonality and its interplay with deterministic components of epidemiological models. Childhood diseases are shown to fall into regions of parameter space of high noise amplification. This type of "endogenous" stochastic resonance may be relevant to population oscillations in nonlinear ecological systems in general.
Figures
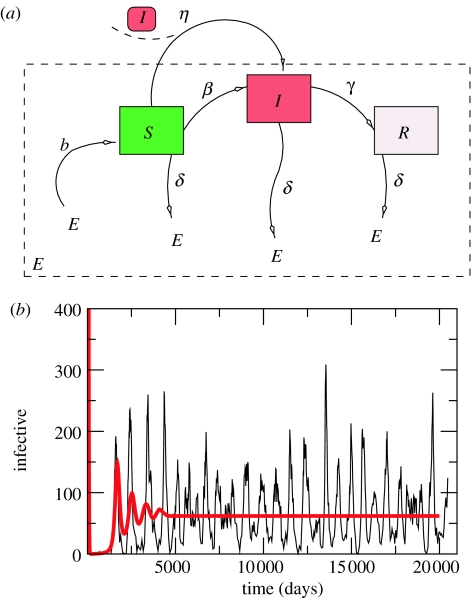
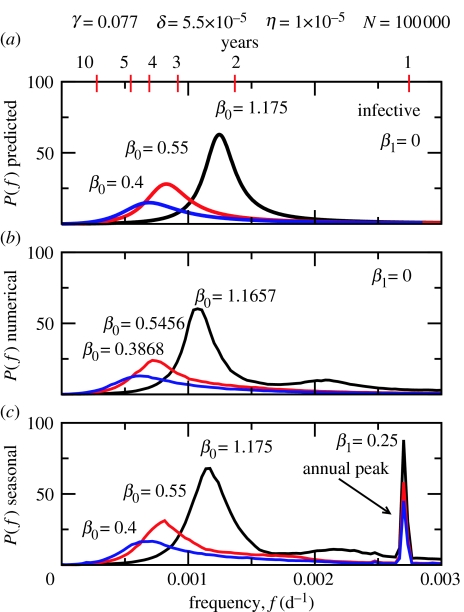
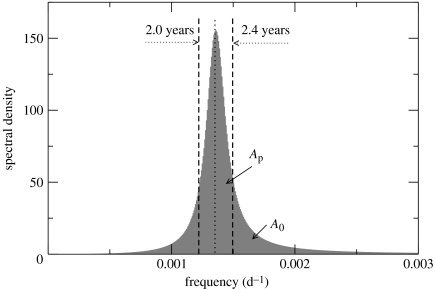
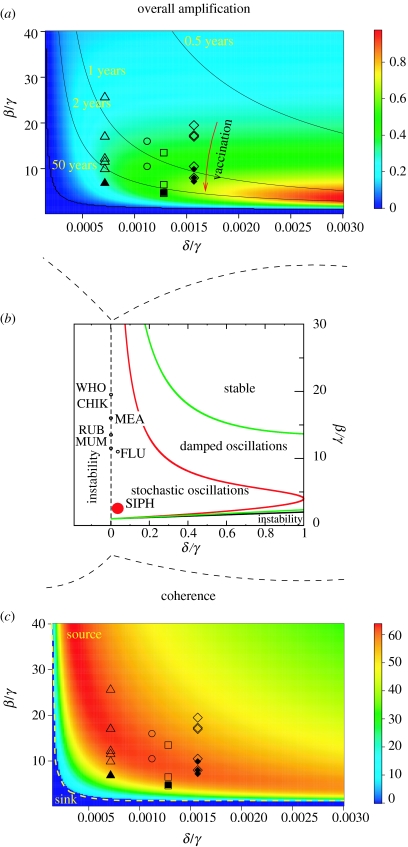
Similar articles
-
Stochastic fluctuations in epidemics on networks.J R Soc Interface. 2008 May 6;5(22):555-66. doi: 10.1098/rsif.2007.1206. J R Soc Interface. 2008. PMID: 17911052 Free PMC article.
-
Regular biennial cycles in epidemics caused by parametric resonance.J Theor Biol. 2017 Feb 21;415:137-144. doi: 10.1016/j.jtbi.2016.12.013. Epub 2016 Dec 20. J Theor Biol. 2017. PMID: 28007555 Free PMC article.
-
Spatial heterogeneity, nonlinear dynamics and chaos in infectious diseases.Stat Methods Med Res. 1995 Jun;4(2):160-83. doi: 10.1177/096228029500400205. Stat Methods Med Res. 1995. PMID: 7582203 Review.
-
Stochastic effects in a seasonally forced epidemic model.Phys Rev E Stat Nonlin Soft Matter Phys. 2010 Oct;82(4 Pt 1):041906. doi: 10.1103/PhysRevE.82.041906. Epub 2010 Oct 6. Phys Rev E Stat Nonlin Soft Matter Phys. 2010. PMID: 21230312
-
Noisy clockwork: time series analysis of population fluctuations in animals.Science. 2001 Jul 27;293(5530):638-43. doi: 10.1126/science.1062226. Science. 2001. PMID: 11474099 Review.
Cited by
-
Dynamics of stochastic epidemics on heterogeneous networks.J Math Biol. 2014 Jun;68(7):1583-605. doi: 10.1007/s00285-013-0679-1. Epub 2013 Apr 30. J Math Biol. 2014. PMID: 23633042
-
Synchrony of sylvatic dengue isolations: a multi-host, multi-vector SIR model of dengue virus transmission in Senegal.PLoS Negl Trop Dis. 2012;6(11):e1928. doi: 10.1371/journal.pntd.0001928. Epub 2012 Nov 29. PLoS Negl Trop Dis. 2012. PMID: 23209867 Free PMC article.
-
Pattern transitions in spatial epidemics: Mechanisms and emergent properties.Phys Life Rev. 2016 Dec;19:43-73. doi: 10.1016/j.plrev.2016.08.002. Epub 2016 Aug 9. Phys Life Rev. 2016. PMID: 27567502 Free PMC article. Review.
-
Fluctuation spectra of large random dynamical systems reveal hidden structure in ecological networks.Nat Commun. 2021 Jun 15;12(1):3625. doi: 10.1038/s41467-021-23757-x. Nat Commun. 2021. PMID: 34131115 Free PMC article.
-
Limits to Causal Inference with State-Space Reconstruction for Infectious Disease.PLoS One. 2016 Dec 28;11(12):e0169050. doi: 10.1371/journal.pone.0169050. eCollection 2016. PLoS One. 2016. PMID: 28030639 Free PMC article.
References
-
- Bartlett M.S. Measles periodicity and community size. J. R. Stat. Soc. A. 1957;120:37–44.
-
- Bauch C.T, Earn D.J.D. Interepidemic intervals in forced and unforced SEIR models. Fields Inst. Commun. 2003a;36:33–44.
-
- Bjørnstad O.N, Finkenstädt B.F, Grenfell B.T. Dynamics of measles epidemics: estimating scaling transmission rates using a time series SIR model. Ecol. Monogr. 2002;72:169–184.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
