

Processing of intronic microRNAs
- PMID: 17255951
- PMCID: PMC1794378
- DOI: 10.1038/sj.emboj.7601512
Processing of intronic microRNAs
Abstract
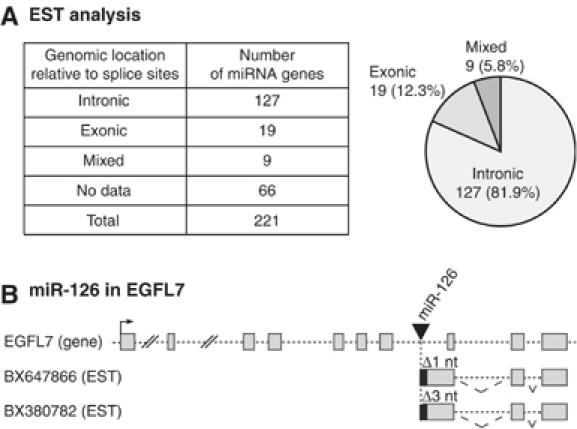
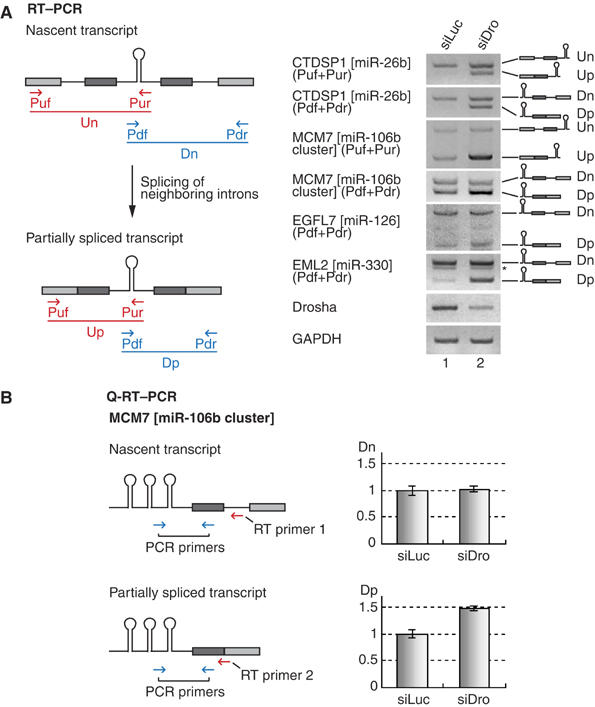
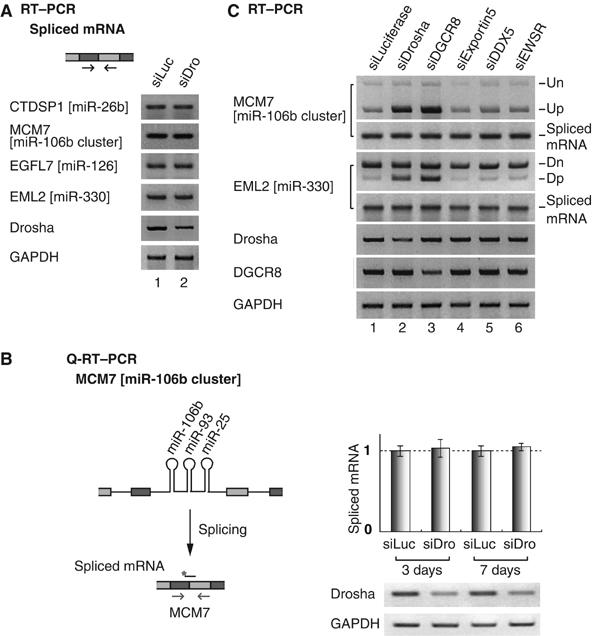
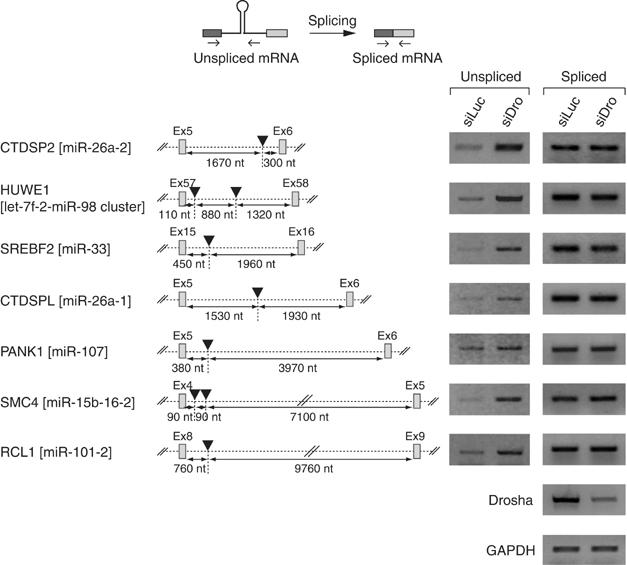
The majority of human microRNA (miRNA) loci are located within intronic regions and are transcribed by RNA polymerase II as part of their hosting transcription units. The primary transcripts are cleaved by Drosha to release approximately 70 nt pre-miRNAs that are subsequently processed by Dicer to generate mature approximately 22 nt miRNAs. It is generally believed that intronic miRNAs are released by Drosha from excised introns after the splicing reaction has occurred. However, our database searches and experiments indicate that intronic miRNAs can be processed from unspliced intronic regions before splicing catalysis. Intriguingly, cleavage of an intron by Drosha does not significantly affect the production of mature mRNA, suggesting that a continuous intron may not be required for splicing and that the exons may be tethered to each other. Hence, Drosha may cleave intronic miRNAs between the splicing commitment step and the excision step, thereby ensuring both miRNA biogenesis and protein synthesis from a single primary transcript. Our study provides a novel example of eukaryotic gene organization and RNA-processing control.
Figures

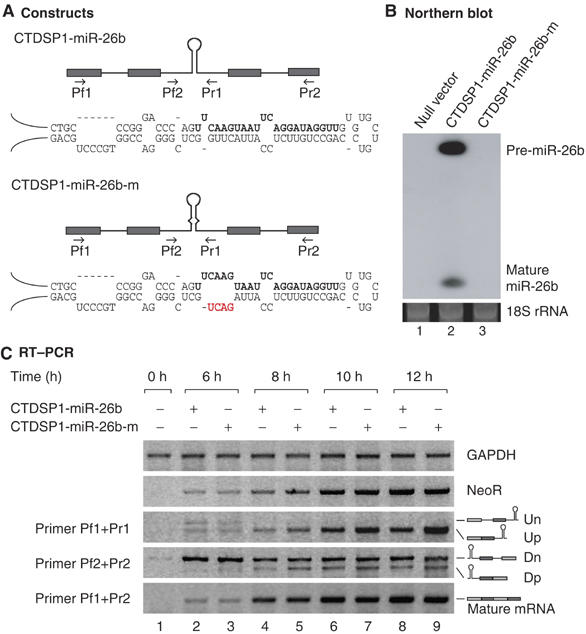
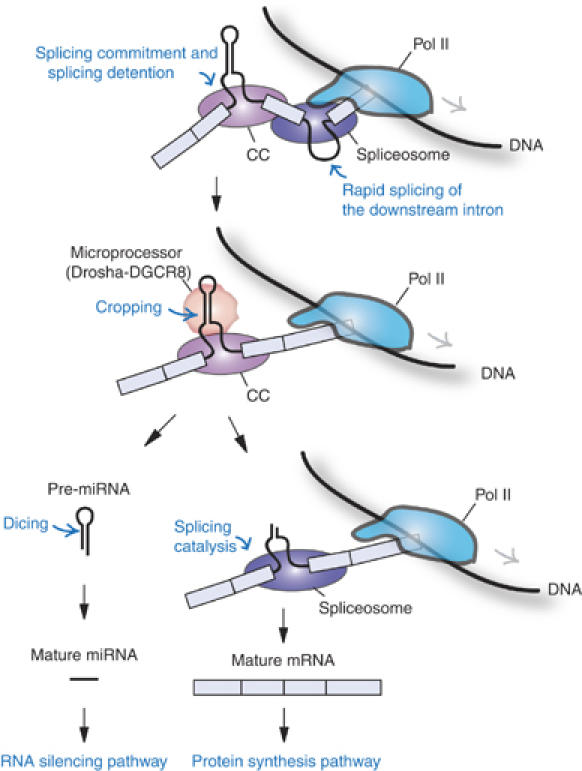
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
