

Adiponectin modulates inflammatory reactions via calreticulin receptor-dependent clearance of early apoptotic bodies
- PMID: 17256056
- PMCID: PMC1770947
- DOI: 10.1172/JCI29709
Adiponectin modulates inflammatory reactions via calreticulin receptor-dependent clearance of early apoptotic bodies
Abstract
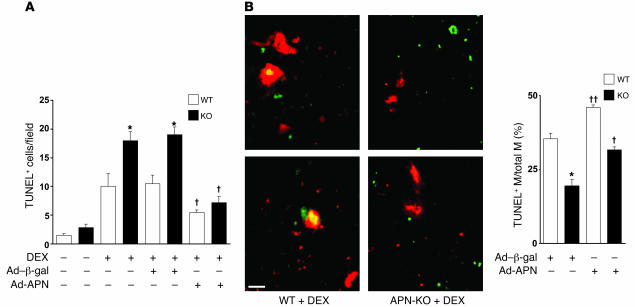
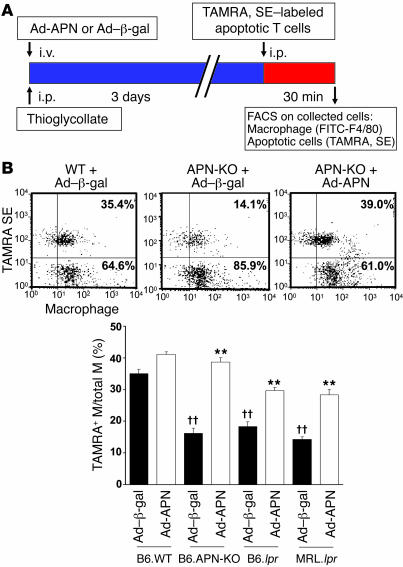
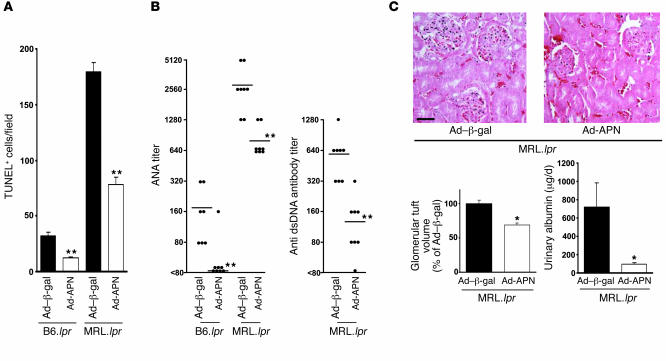
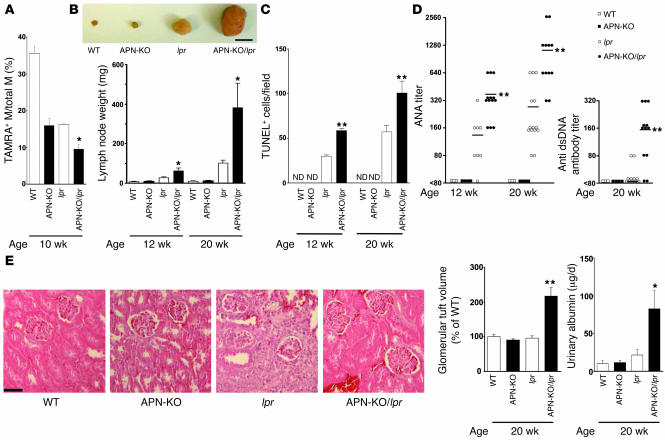
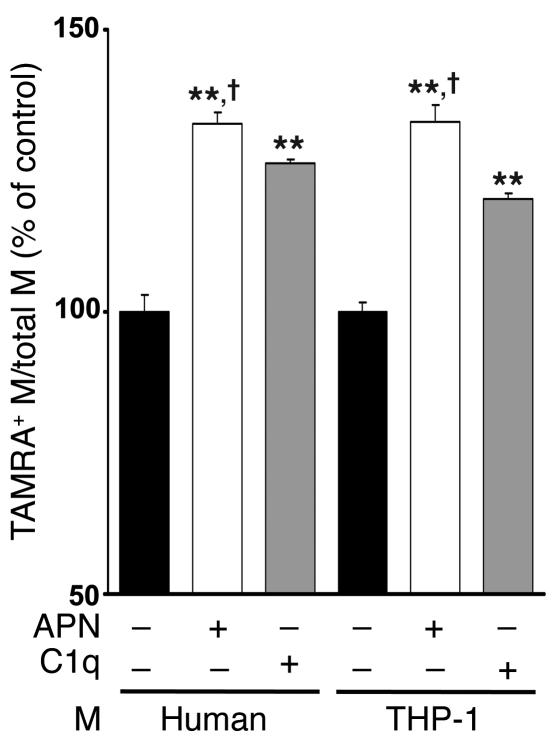
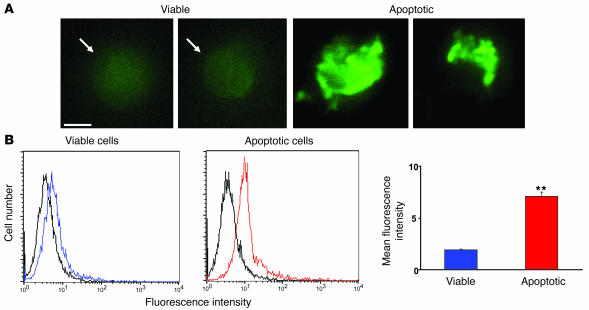
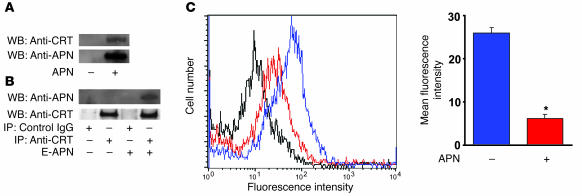
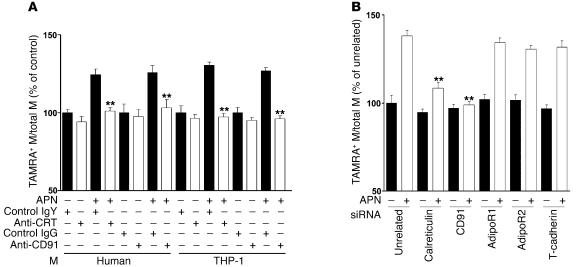
Obesity and type 2 diabetes are associated with chronic inflammation. Adiponectin is an adipocyte-derived hormone with antidiabetic and antiinflammatory actions. Here, we demonstrate what we believe to be a previously undocumented activity of adiponectin, facilitating the uptake of early apoptotic cells by macrophages, an essential feature of immune system function. Adiponectin-deficient (APN-KO) mice were impaired in their ability to clear apoptotic thymocytes in response to dexamethasone treatment, and these animals displayed a reduced ability to clear early apoptotic cells that were injected into their intraperitoneal cavities. Conversely, adiponectin administration promoted the clearance of apoptotic cells by macrophages in both APN-KO and wild-type mice. Adiponectin overexpression also promoted apoptotic cell clearance and reduced features of autoimmunity in lpr mice whereas adiponectin deficiency in lpr mice led to a further reduction in apoptotic cell clearance, which was accompanied by exacerbated systemic inflammation. Adiponectin was capable of opsonizing apoptotic cells, and phagocytosis of cell corpses was mediated by the binding of adiponectin to calreticulin on the macrophage cell surface. We propose that adiponectin protects the organism from systemic inflammation by promoting the clearance of early apoptotic cells by macrophages through a receptor-dependent pathway involving calreticulin.
Figures
References
-
- Ruan H., Lodish H.F. Regulation of insulin sensitivity by adipose tissue-derived hormones and inflammatory cytokines. Curr. Opin. Lipidol. 2004;15:297–302. - PubMed
-
- Spiegelman B.M., Choy L., Hotamisligil G.S., Graves R.A., Tontonoz P. Regulation of adipocyte gene expression in differentiation and syndromes of obesity/diabetes. J. Biol. Chem. 1993;268:6823–6826. - PubMed
-
- Maeda K., et al. cDNA cloning and expression of a novel adipose specific collagen-like factor, apM1 (AdiPose Most abundant Gene transcript 1). Biochem. Biophys. Res. Commun. 1996;221:286–289. - PubMed
-
- Hu E., Liang P., Spiegelman B.M.1996. . AdipoQ is a novel adipose-specific gene dysregulated in obesity. J. Biol. Chem. 27110697–10703. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
