

Dimer dissociation and unfolding mechanism of coagulation factor XI apple 4 domain: spectroscopic and mutational analysis
- PMID: 17257616
- PMCID: PMC1945241
- DOI: 10.1016/j.jmb.2006.12.066
Dimer dissociation and unfolding mechanism of coagulation factor XI apple 4 domain: spectroscopic and mutational analysis
Abstract
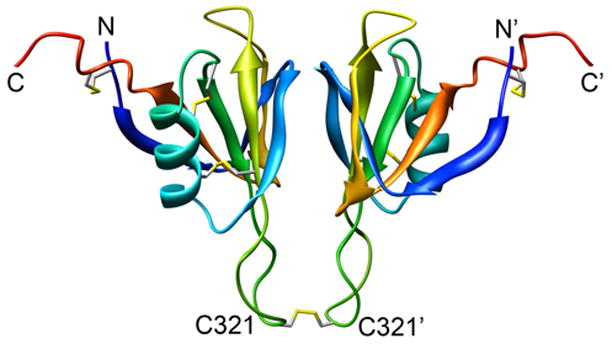
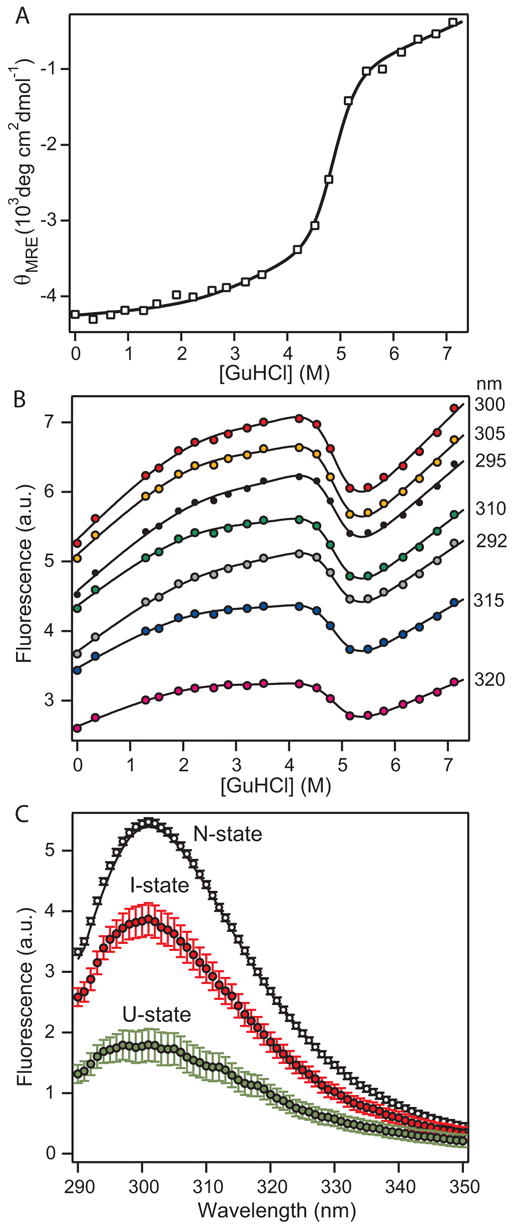
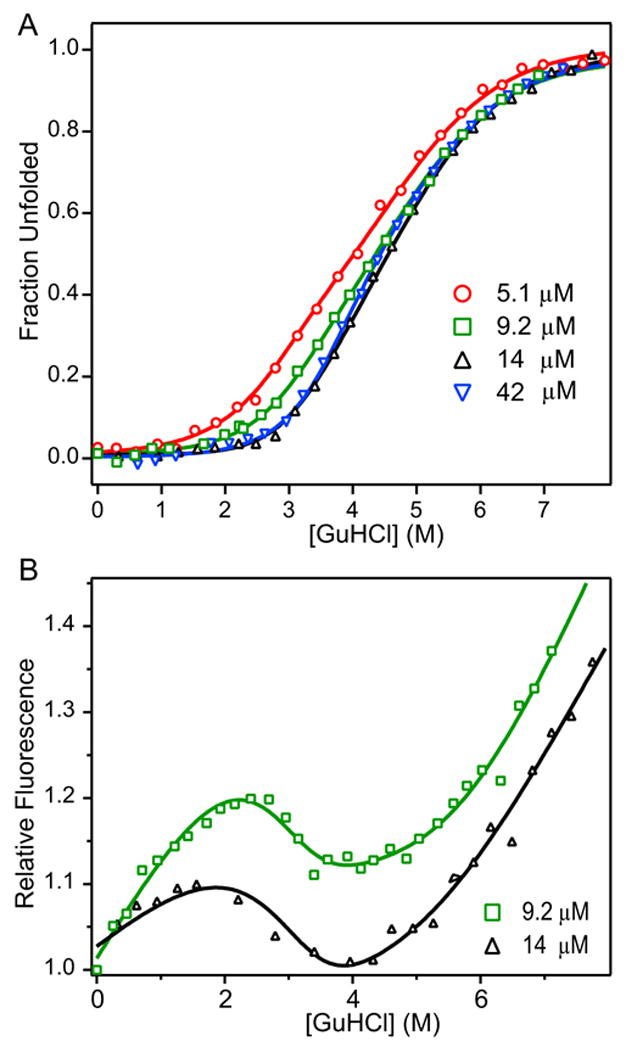
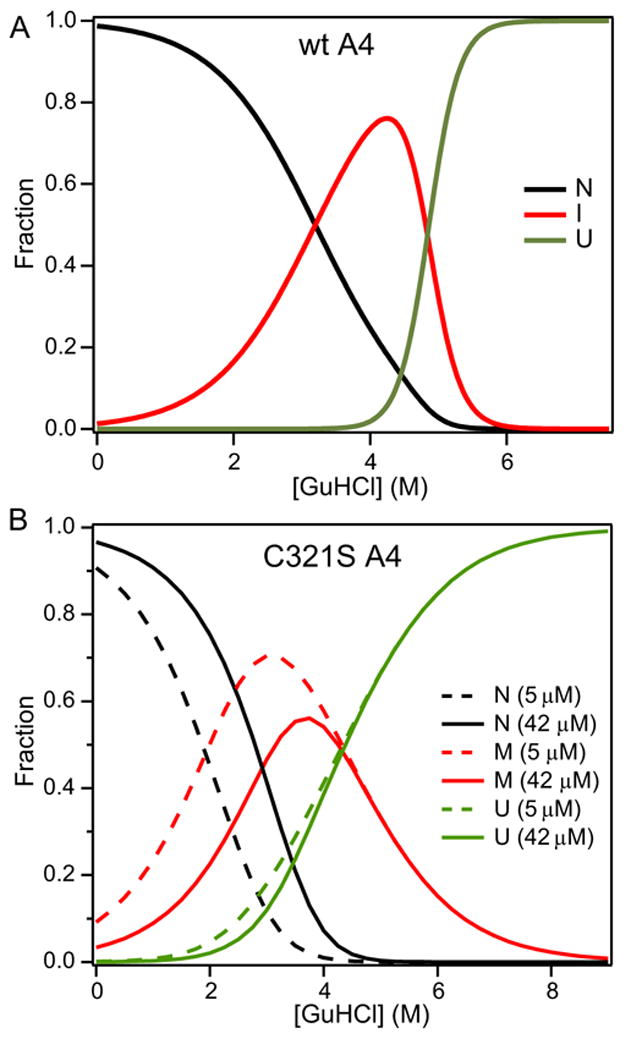
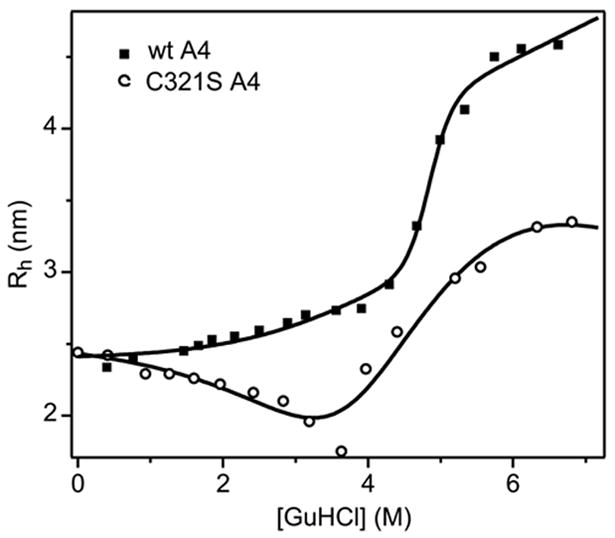
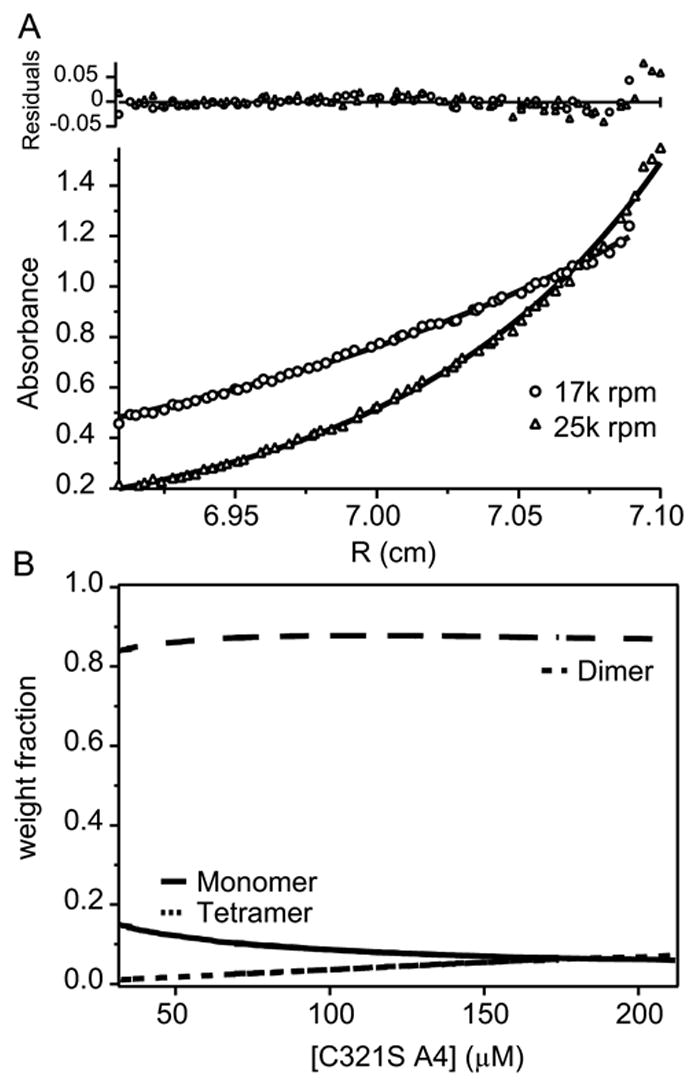
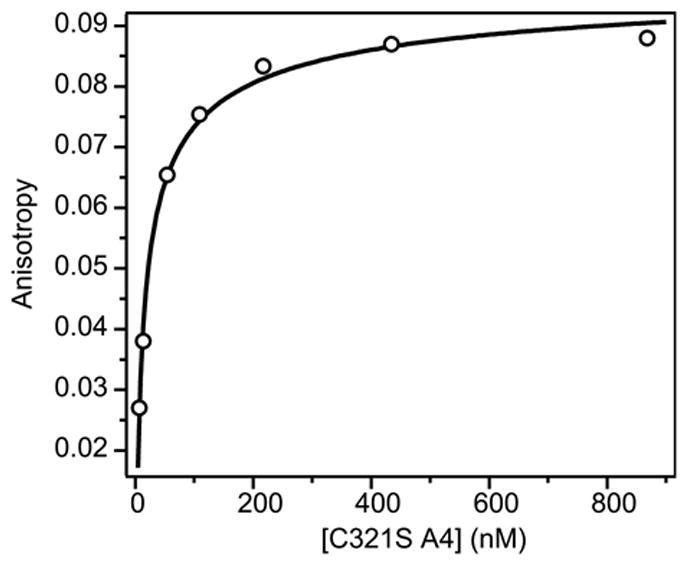
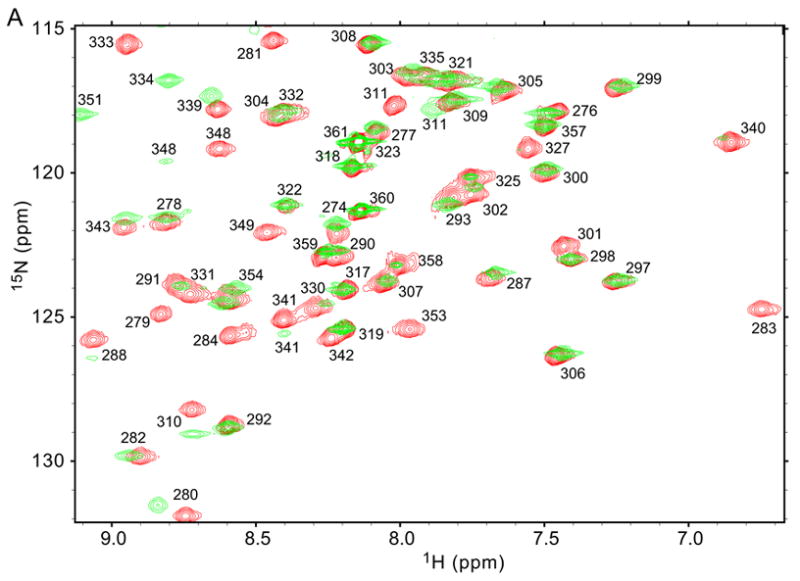
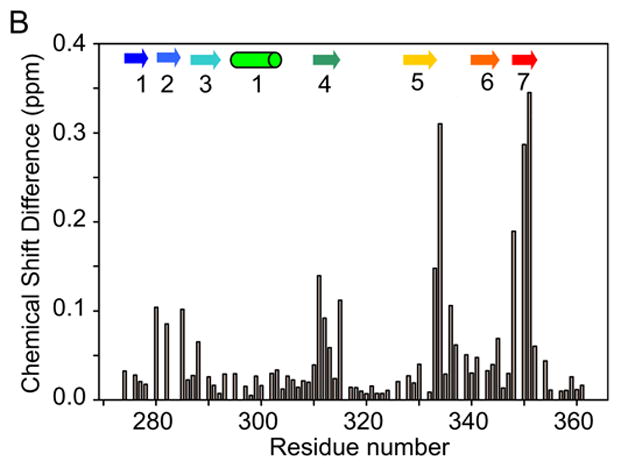
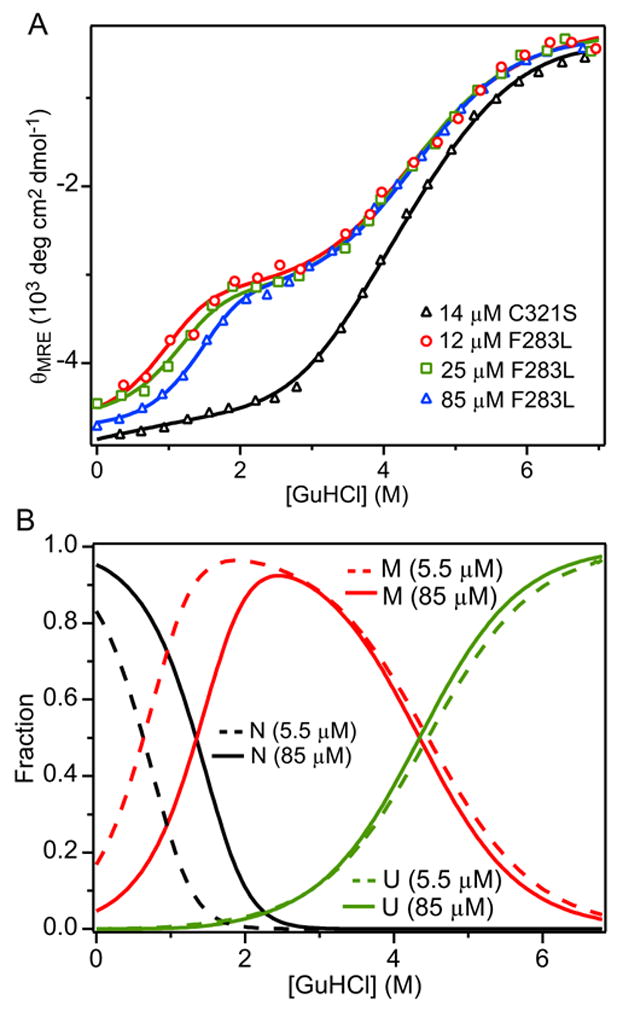
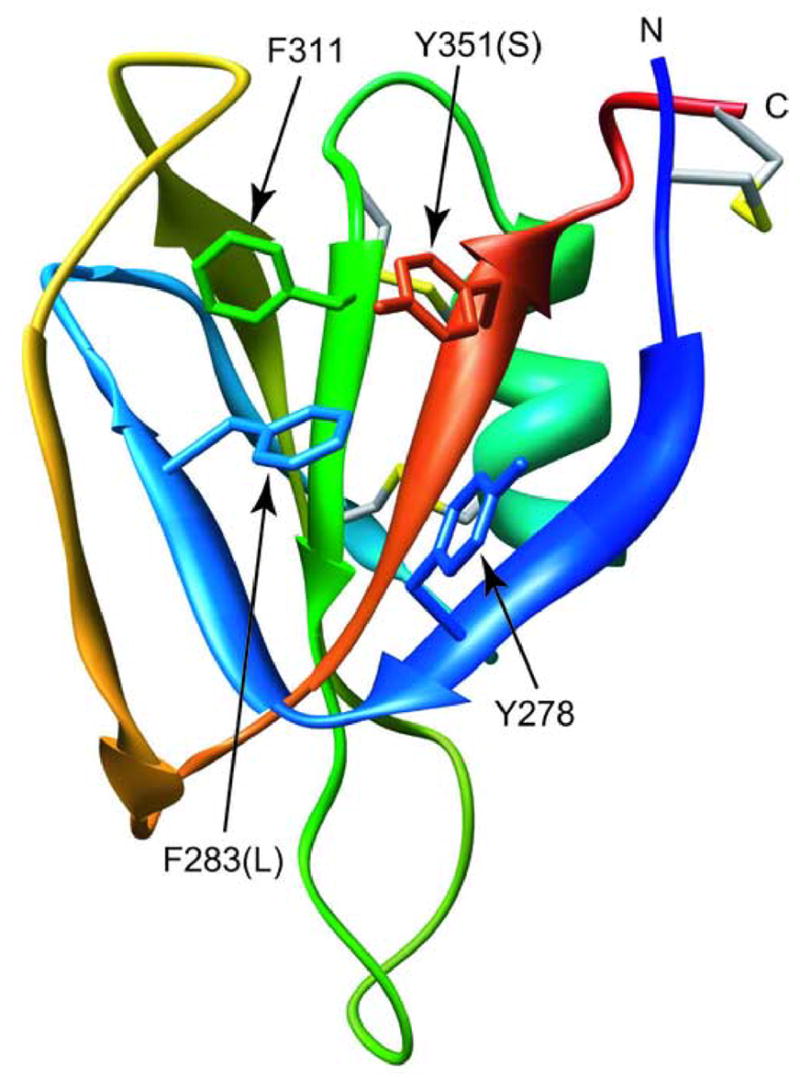
The blood coagulation protein factor XI (FXI) consists of a pair of disulfide-linked chains each containing four apple domains and a catalytic domain. The apple 4 domain (A4; F272-E362) mediates non-covalent homodimer formation even when the cysteine involved in an intersubunit disulfide is mutated to serine (C321S). To understand the role of non-covalent interactions stabilizing the FXI dimer, equilibrium unfolding of wild-type A4 and its C321S variant was monitored by circular dichroism, intrinsic tyrosine fluorescence and dynamic light scattering measurements as a function of guanidine hydrochloride concentration. Global analysis of the unimolecular unfolding transition of wild-type A4 revealed a partially unfolded equilibrium intermediate at low to moderate denaturant concentrations. The optically detected equilibrium of C321S A4 also fits best to a three-state model in which the native dimer unfolds via a monomeric intermediate state. Dimer dissociation is characterized by a dissociation constant, K(d), of approximately 90 nM (in terms of monomer), which is in agreement with the dissociation constant measured independently using fluorescence anisotropy. The results imply that FXI folding occurs via a monomeric equilibrium intermediate. This observation sheds light on the effect of certain naturally occurring mutations, such as F283L, which lead to intracellular accumulation of non-native forms of FXI. To investigate the structural and energetic consequences of the F283L mutation, which perturbs a cluster of aromatic side-chains within the core of the A4 monomer, it was introduced into the dissociable dimer, C321S A4. NMR chemical shift analysis confirmed that the mutant can assume a native-like dimeric structure. However, equilibrium unfolding measurements show that the mutation causes a fourfold increase in the K(d) value for dissociation of the native dimer and a 1 kcal/mol stabilization of the monomer, resulting in a highly populated intermediate. Since the F283 side-chain does not directly participate in the dimer interface, we propose that the F283L mutation leads to increased dimer dissociation by stabilizing a monomeric state with altered side-chain packing that is unfavorable for homodimer formation.
Figures
References
-
- Walsh PN, Gailani D. Factor XI. In: Colman RW, Hirsh J, Marder VJ, Clowes AW, George JN, editors. Hemostasis and Thrombosis: Basic Principles and Clinical Practice. 5. Chapter 12. Lippincott Williams & Wilkins; Philadelphia: 2006.
-
- Fujikawa K, Legaz ME, Kato H, Davie EW. The mechanism of activation of bovine factor IX (Christmas factor) by bovine factor XIa (activated plasma thromboplastin antecedent) Biochemistry. 1974;13:4508–16. - PubMed
-
- Gailani D, Broze GJ., Jr Factor XI activation in a revised model of blood coagulation. Science. 1991;253:909–12. - PubMed
-
- Naito K, Fujikawa K. Activation of human blood coagulation factor XI independent of factor XII. Factor XI is activated by thrombin and factor XIa in the presence of negatively charged surfaces. J Biol Chem. 1991;266:7353–8. - PubMed
-
- Papagrigoriou E, McEwan PA, Walsh PN, Emsley J. Crystal structure of the factor XI zymogen reveals a pathway for transactivation. Nat Struct Mol Biol. 2006;13:557–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
