

Disruption of a sugar transporter gene cluster in a hyperthermophilic archaeon using a host-marker system based on antibiotic resistance
- PMID: 17259314
- PMCID: PMC1855824
- DOI: 10.1128/JB.01692-06
Disruption of a sugar transporter gene cluster in a hyperthermophilic archaeon using a host-marker system based on antibiotic resistance
Abstract
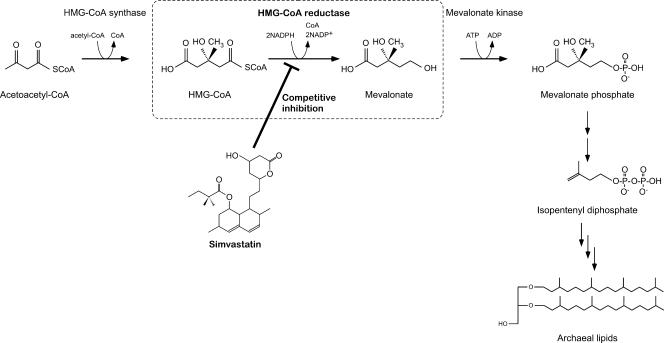
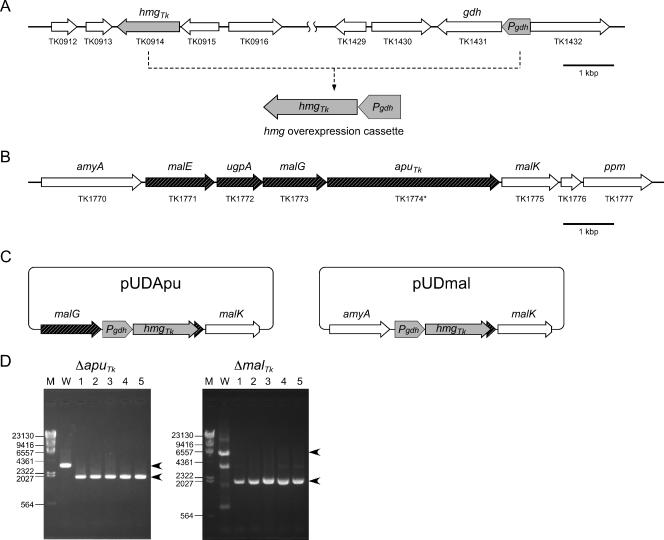
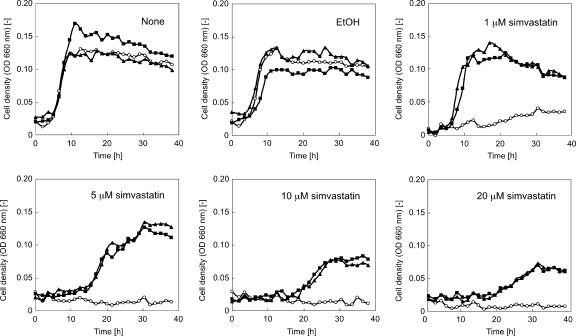
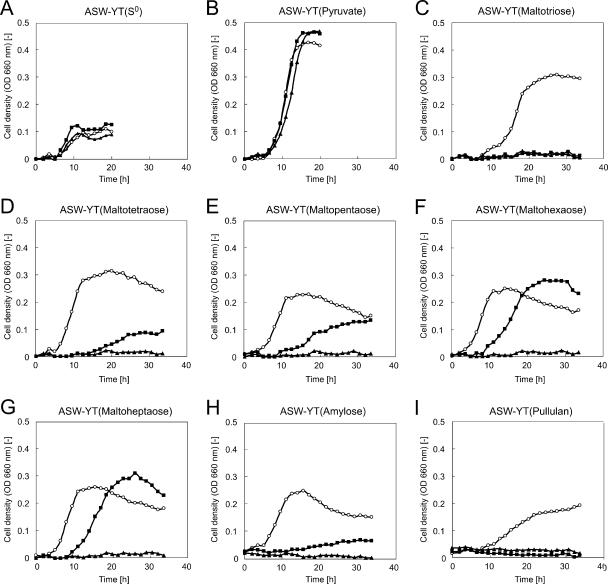
We have developed a gene disruption system in the hyperthermophilic archaeon Thermococcus kodakaraensis using the antibiotic simvastatin and a fusion gene designed to overexpress the 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA) reductase gene (hmg(Tk)) with the glutamate dehydrogenase promoter. With this system, we disrupted the T. kodakaraensis amylopullulanase gene (apu(Tk)) or a gene cluster which includes apu(Tk) and genes encoding components of a putative sugar transporter. Disruption plasmids were introduced into wild-type T. kodakaraensis KOD1 cells, and transformants exhibiting resistance to 4 microM simvastatin were isolated. The transformants exhibited growth in the presence of 20 microM simvastatin, and we observed a 30-fold increase in intracellular HMG-CoA reductase activity. The expected gene disruption via double-crossover recombination occurred at the target locus, but we also observed recombination events at the hmg(Tk) locus when the endogenous hmg(Tk) gene was used. This could be avoided by using the corresponding gene from Pyrococcus furiosus (hmg(Pf)) or by linearizing the plasmid prior to transformation. While both gene disruption strains displayed normal growth on amino acids or pyruvate, cells without the sugar transporter genes could not grow on maltooligosaccharides or polysaccharides, indicating that the gene cluster encodes the only sugar transporter involved in the uptake of these compounds. The Deltaapu(Tk) strain could not grow on pullulan and displayed only low levels of growth on amylose, suggesting that Apu(Tk) is a major polysaccharide-degrading enzyme in T. kodakaraensis.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
