

LARK activates posttranscriptional expression of an essential mammalian clock protein, PERIOD1
- PMID: 17264215
- PMCID: PMC1794262
- DOI: 10.1073/pnas.0607567104
LARK activates posttranscriptional expression of an essential mammalian clock protein, PERIOD1
Abstract
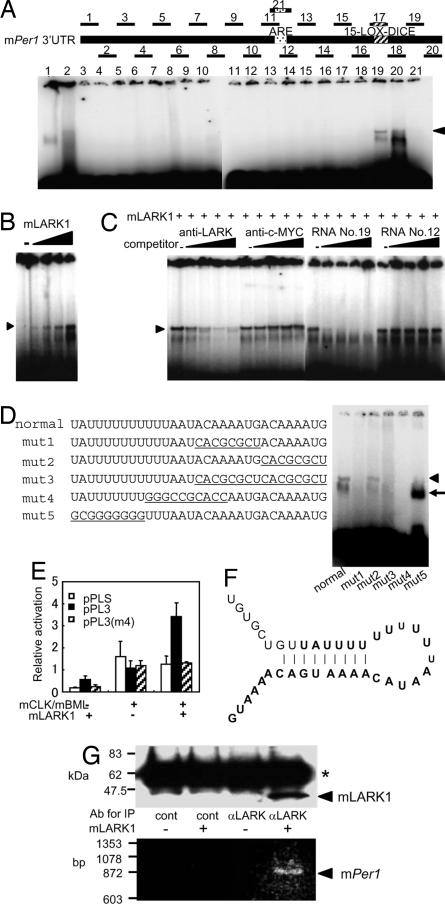
The mammalian molecular clock is composed of feedback loops to keep circadian 24-h rhythms. Although much focus has been on transcriptional regulation, it is clear that posttranscriptional controls also play important roles in molecular circadian clocks. In this study, we found that mouse LARK (mLARK), an RNA binding protein, activates the posttranscriptional expression of the mouse Period1 (mPer1) mRNA. A strong circadian cycling of the mLARK protein is observed in the suprachiasmatic nuclei with a phase similar to that of mPER1, although the level of the Lark transcripts are not rhythmic. We demonstrate that LARK causes increased mPER1 protein levels, most likely through translational regulation and that the LARK1 protein binds directly to a cis element in the 3' UTR of the mPer1 mRNA. Alterations of mLark expression in cycling cells caused significant changes in circadian period, with mLark knockdown by siRNA resulting in a shorter circadian period, and the overexpression of mLARK1 resulting in a lengthened period. These data indicate that mLARKs are novel posttranscriptional regulators of mammalian circadian clocks.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





Similar articles
-
Expression of mPer1 and mPer2, two mammalian clock genes, in murine bone marrow.Biochem Biophys Res Commun. 2000 Sep 24;276(2):724-8. doi: 10.1006/bbrc.2000.3536. Biochem Biophys Res Commun. 2000. PMID: 11027538
-
Long-term constant light induces constitutive elevated expression of mPER2 protein in the murine SCN: a molecular basis for Aschoff's rule?J Biol Rhythms. 2005 Feb;20(1):3-14. doi: 10.1177/0748730404272858. J Biol Rhythms. 2005. PMID: 15654066
-
Circadian and photic regulation of clock and clock-controlled proteins in the suprachiasmatic nuclei of calorie-restricted mice.Eur J Neurosci. 2007 Jun;25(12):3691-701. doi: 10.1111/j.1460-9568.2007.05626.x. Eur J Neurosci. 2007. PMID: 17610588
-
Posttranscriptional regulation of mammalian circadian clock output.Cold Spring Harb Symp Quant Biol. 2007;72:145-56. doi: 10.1101/sqb.2007.72.022. Cold Spring Harb Symp Quant Biol. 2007. PMID: 18419272 Review.
-
Role of phosphorylation in the mammalian circadian clock.Cold Spring Harb Symp Quant Biol. 2007;72:167-76. doi: 10.1101/sqb.2007.72.036. Cold Spring Harb Symp Quant Biol. 2007. PMID: 18419274 Review.
Cited by
-
The distinct roles of zinc finger CCHC-type (ZCCHC) superfamily proteins in the regulation of RNA metabolism.RNA Biol. 2021 Dec;18(12):2107-2126. doi: 10.1080/15476286.2021.1909320. Epub 2021 May 4. RNA Biol. 2021. PMID: 33787465 Free PMC article. Review.
-
New insights into non-transcriptional regulation of mammalian core clock proteins.J Cell Sci. 2020 Sep 15;133(18):jcs241174. doi: 10.1242/jcs.241174. J Cell Sci. 2020. PMID: 32934011 Free PMC article. Review.
-
Cold-induced RNA-binding proteins regulate circadian gene expression by controlling alternative polyadenylation.Sci Rep. 2013;3:2054. doi: 10.1038/srep02054. Sci Rep. 2013. PMID: 23792593 Free PMC article.
-
MicroRNA-185 oscillation controls circadian amplitude of mouse Cryptochrome 1 via translational regulation.Mol Biol Cell. 2013 Jul;24(14):2248-55. doi: 10.1091/mbc.E12-12-0849. Epub 2013 May 22. Mol Biol Cell. 2013. PMID: 23699394 Free PMC article.
-
Translational profiling of clock cells reveals circadianly synchronized protein synthesis.PLoS Biol. 2013 Nov;11(11):e1001703. doi: 10.1371/journal.pbio.1001703. Epub 2013 Nov 5. PLoS Biol. 2013. PMID: 24348200 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
