

Quest for the basic plan of nervous system circuitry
- PMID: 17267046
- PMCID: PMC2243186
- DOI: 10.1016/j.brainresrev.2006.12.006
Quest for the basic plan of nervous system circuitry
Abstract
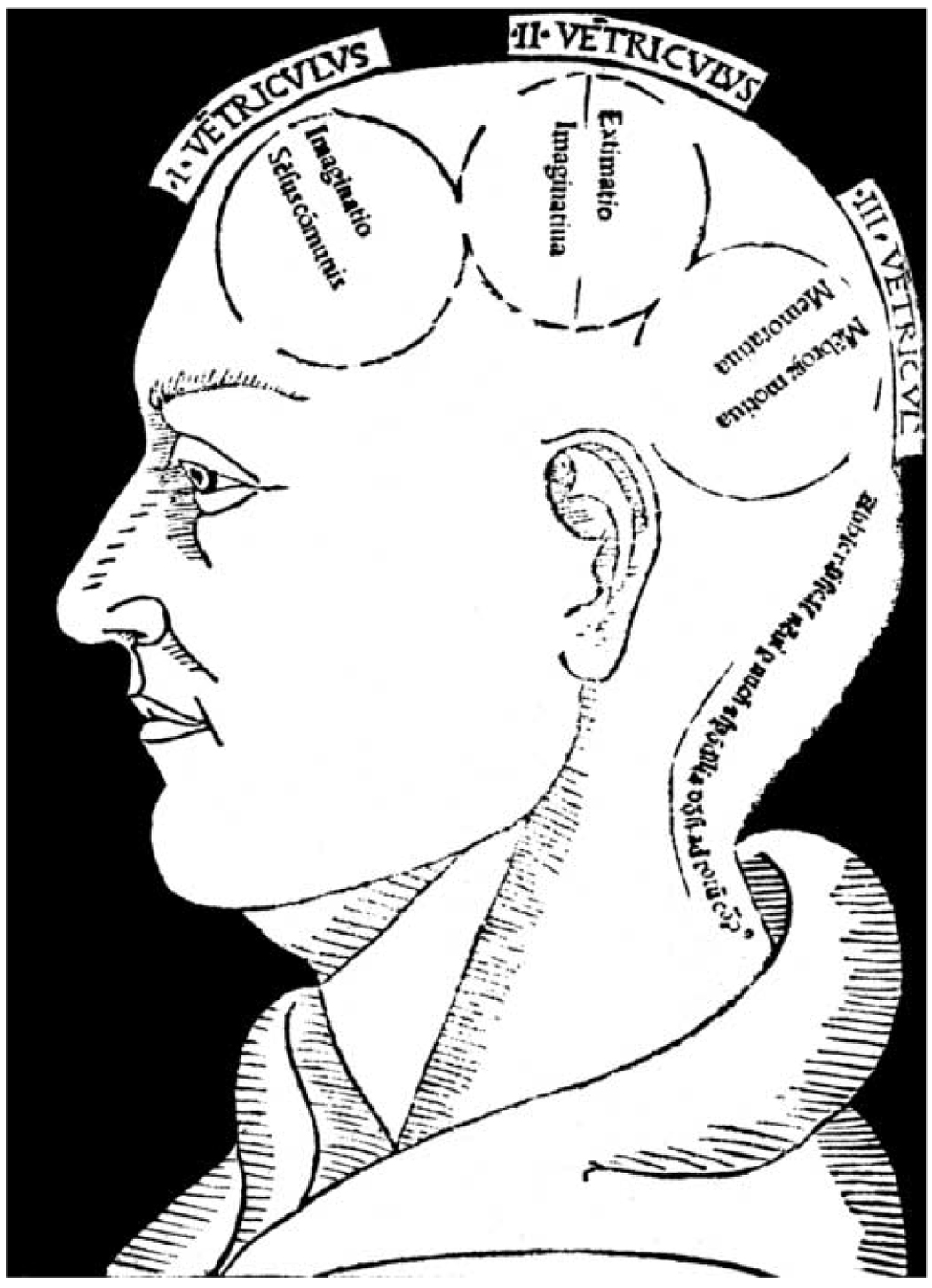
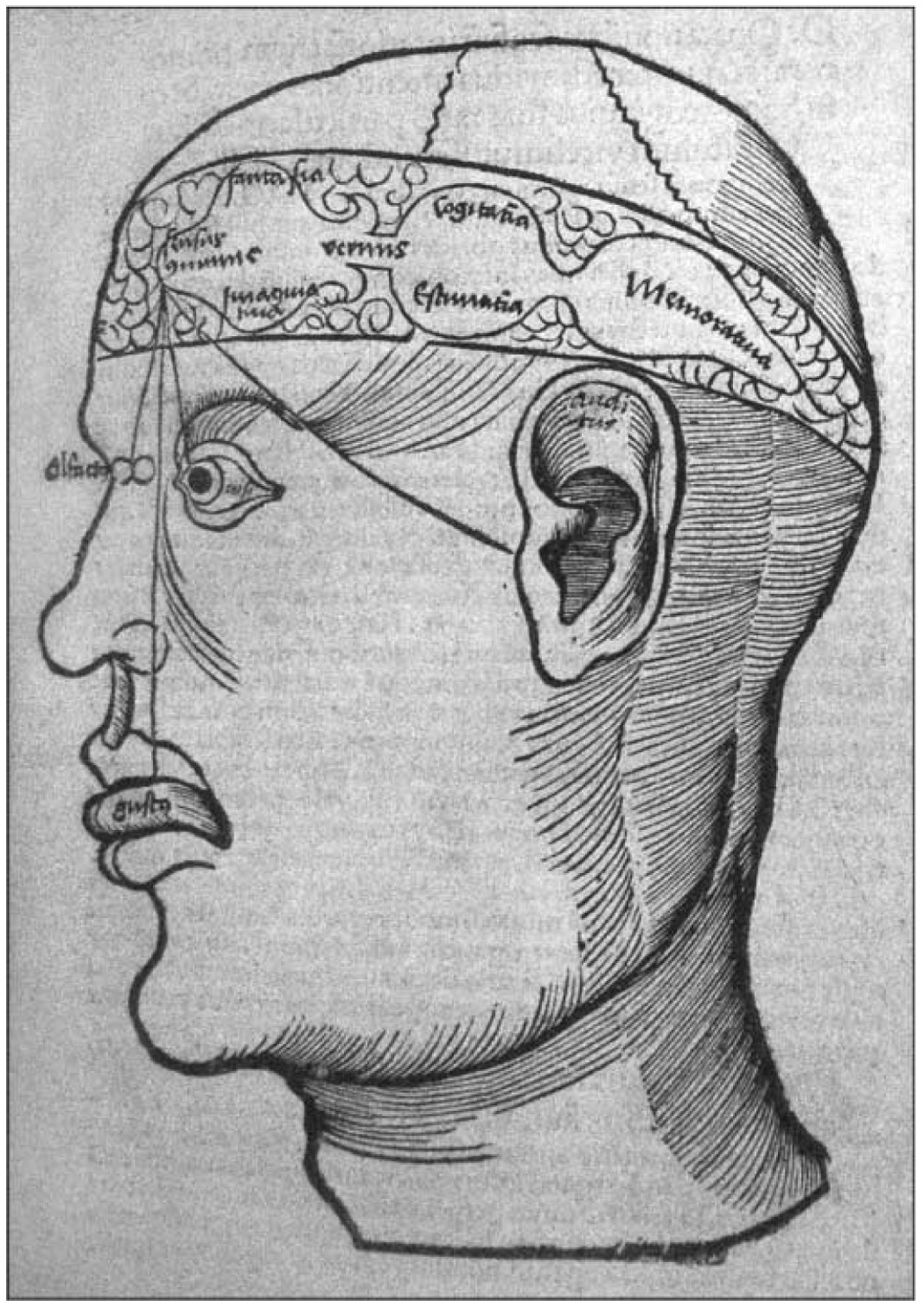
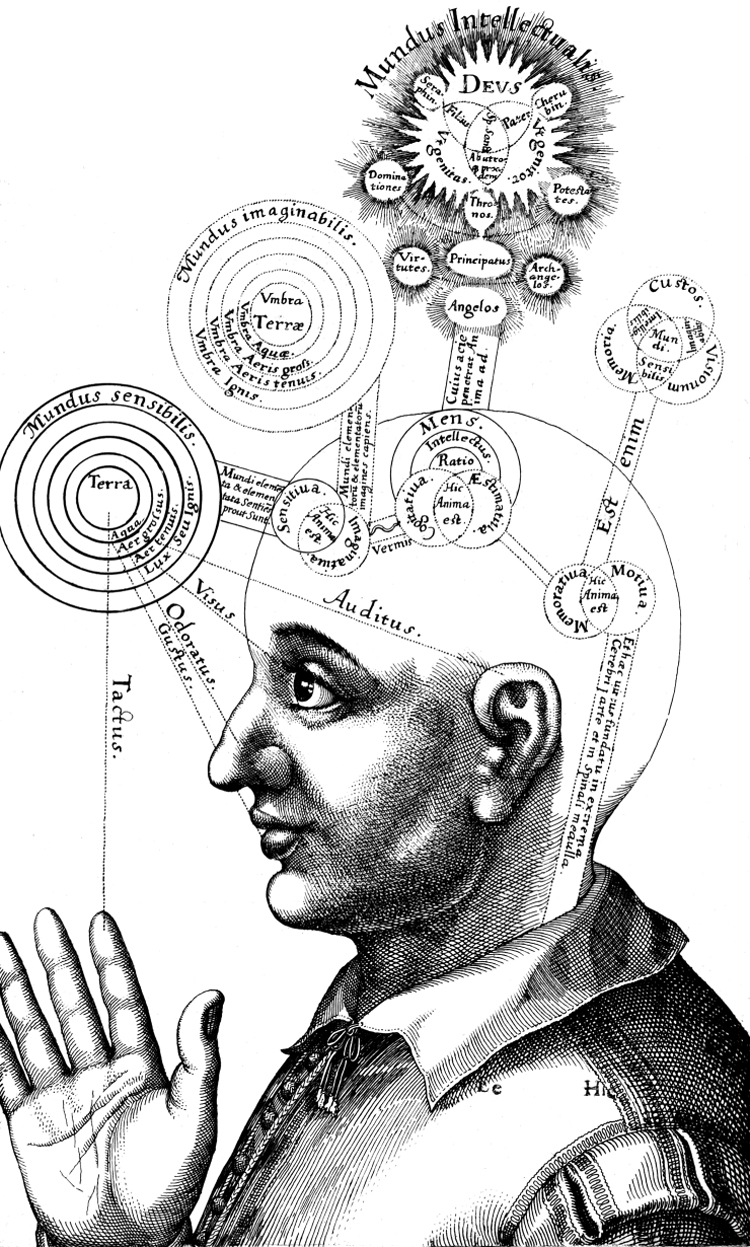
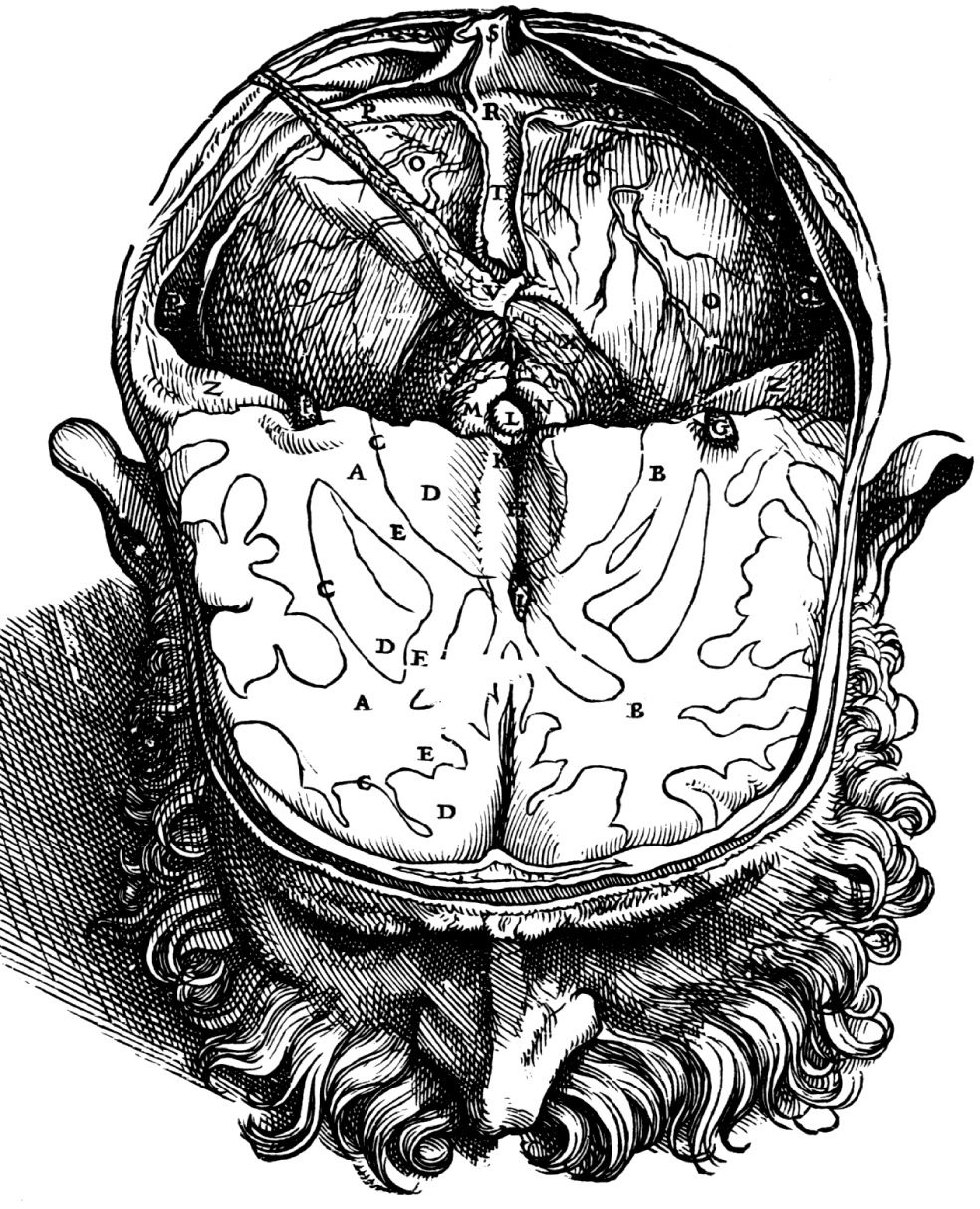
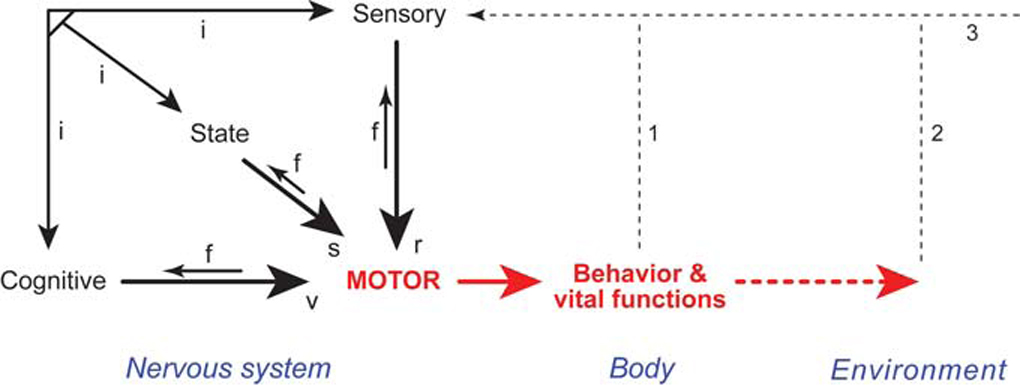
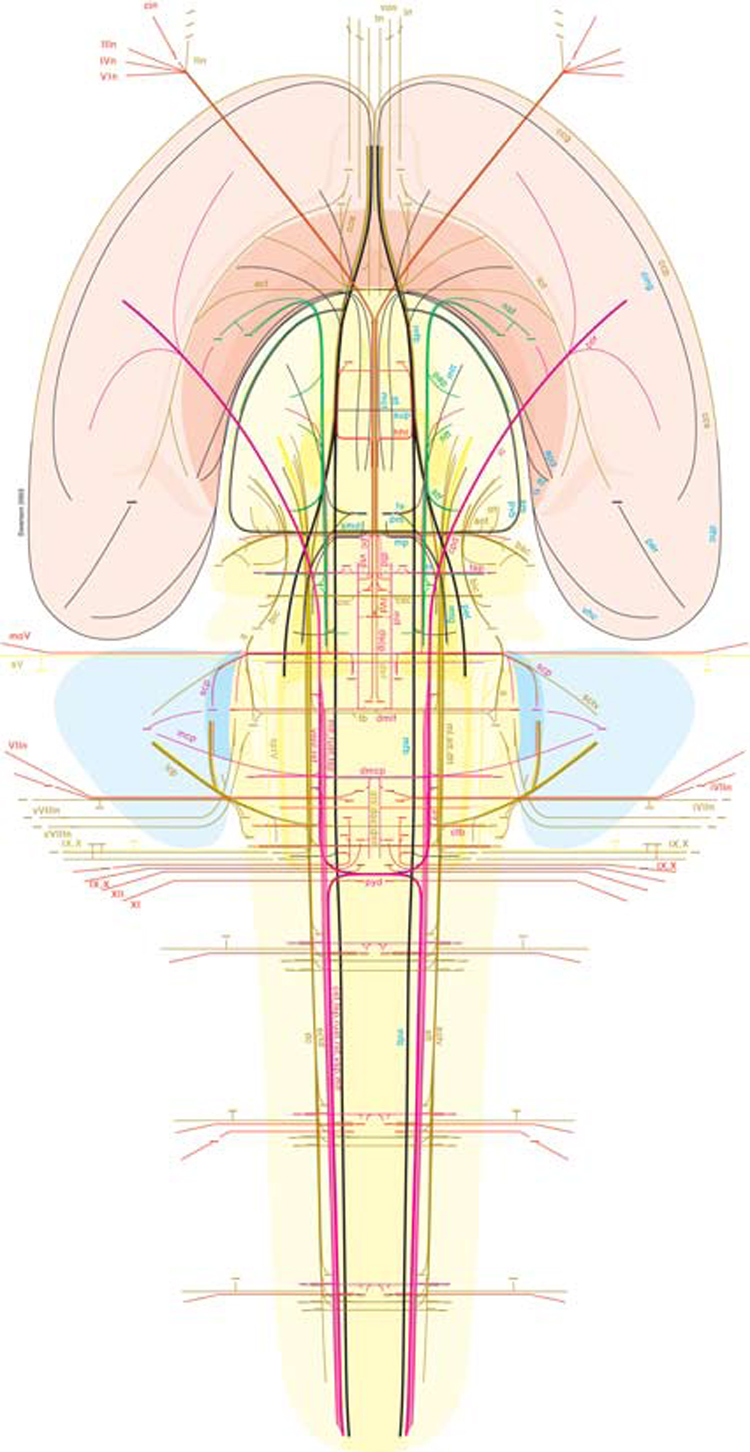
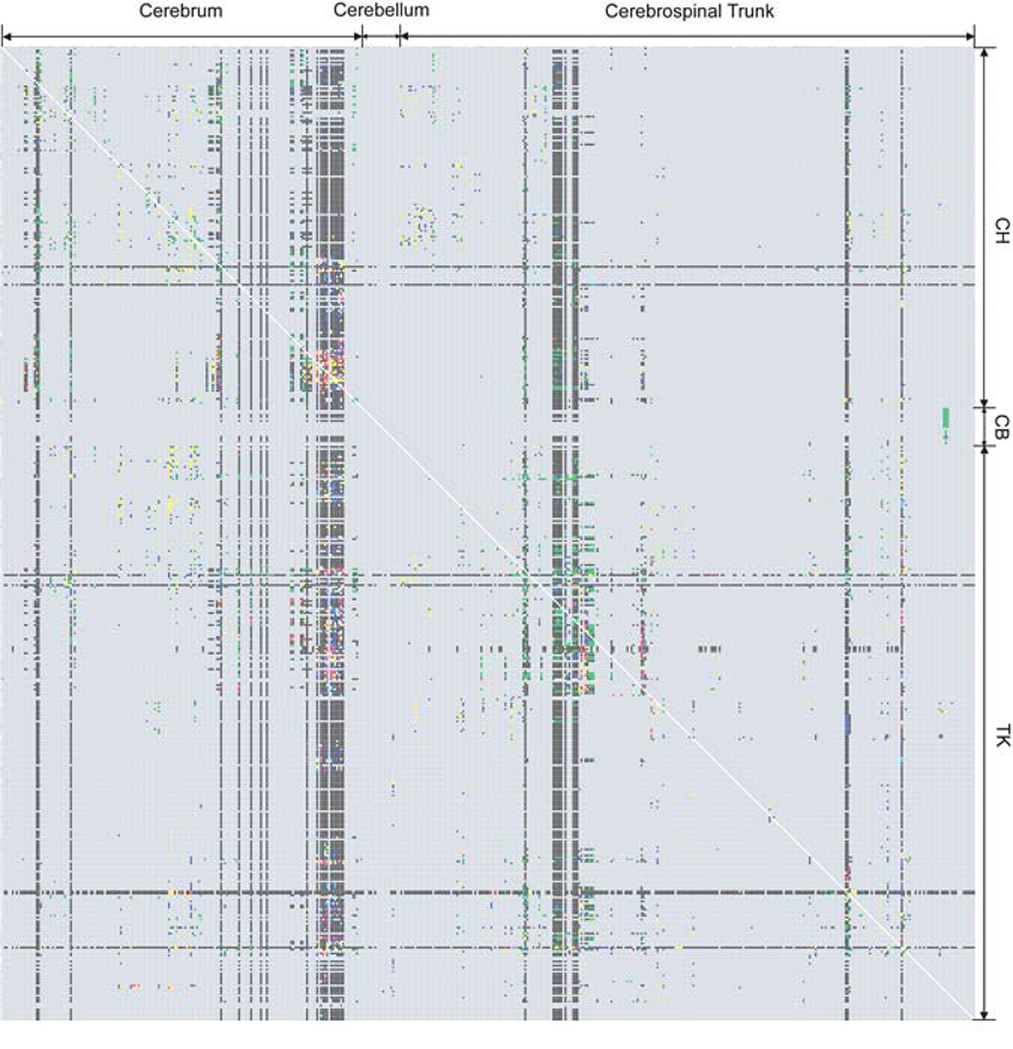
The basic plan of nervous system organization has been investigated since classical antiquity. The first model centered on pneumas pumped from sensory nerves through the ventricular system and out motor nerves to muscles. It was popular well into the 17th century and diverted attention from the organization of brain parenchyma itself. Willis focused on gray matter production and white matter conduction of pneumas in 1664, and by the late 19th century a clear cellular model of nervous system organization based on sensory, motor, and association neuron classes transmitting nerve impulses was elaborated by Cajal and his contemporaries. Today, revolutionary advances in experimental pathway tracing methods, molecular genetics, and computer science inspire systems neuroscience. Seven minimal requirements are outlined for knowledge management systems capable of describing, analyzing, and modeling the basic plan of nervous system circuitry in general, and the plan evolved for vertebrates, for mammals, and ultimately for humans in particular. The goal remains a relatively simple, easy to understand model analogous to the one Harvey elaborated in 1628 for blood circulation in the cardiovascular system. As Cajal wrote in 1909, "To extend our understanding of neural function to the most complex human physiological and psychological activities, it is essential that we first generate a clear and accurate view of the structure of the relevant centers, and of the human brain itself, so that the basic plan--the overview--can be grasped in the blink of an eye."
Figures
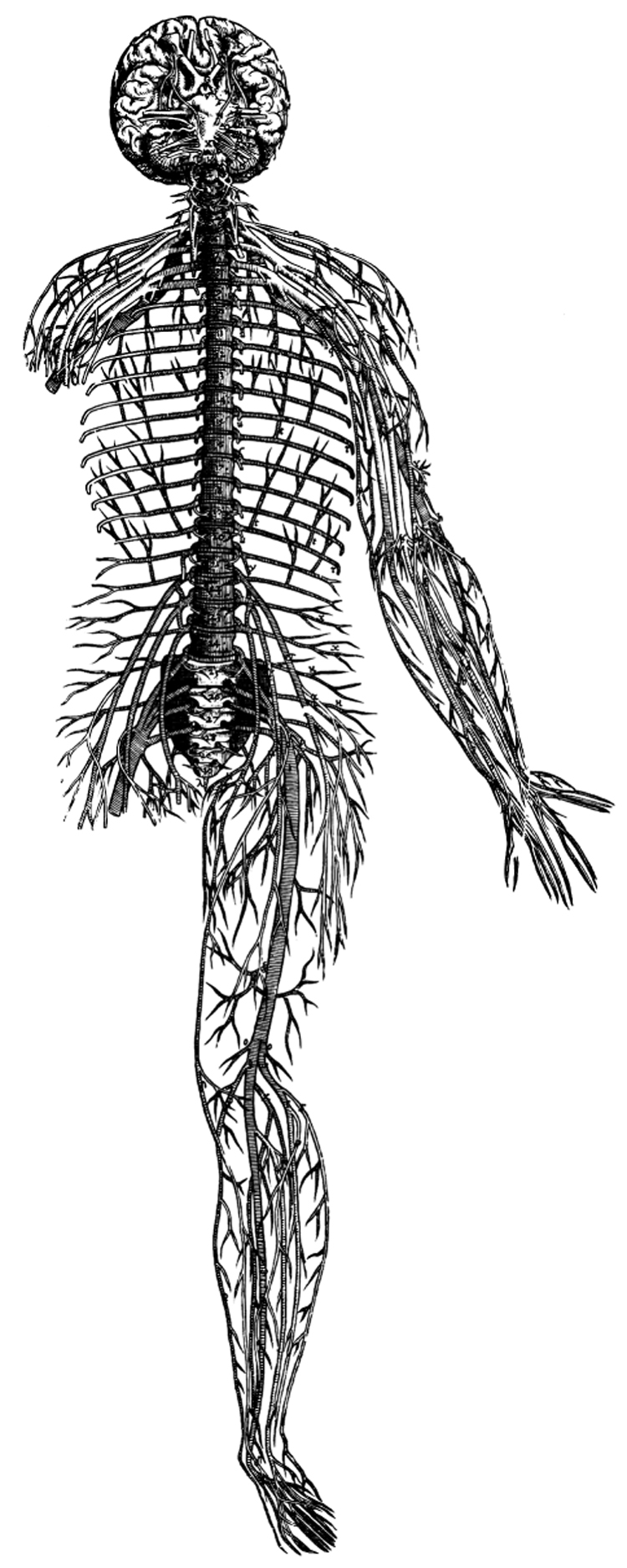
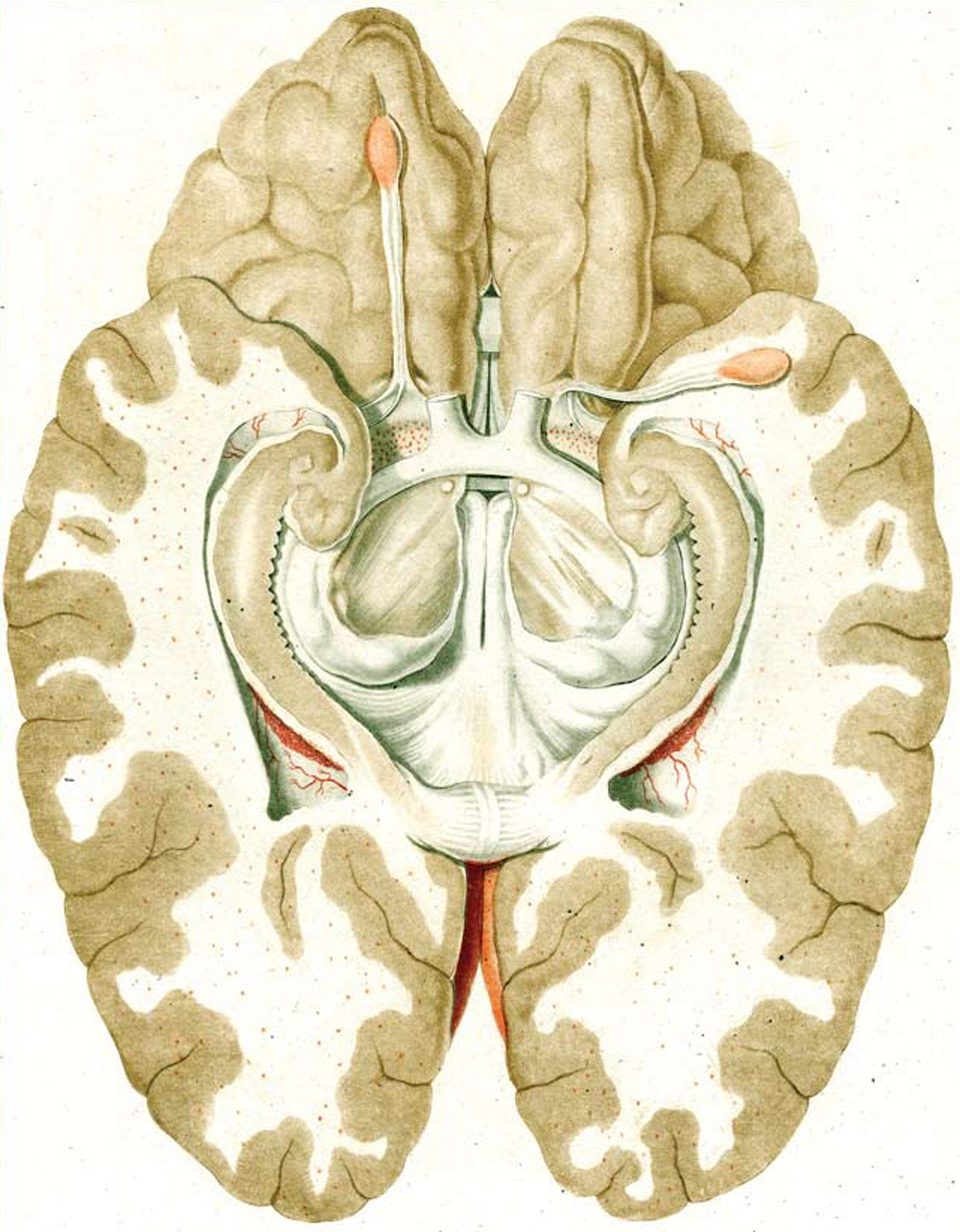
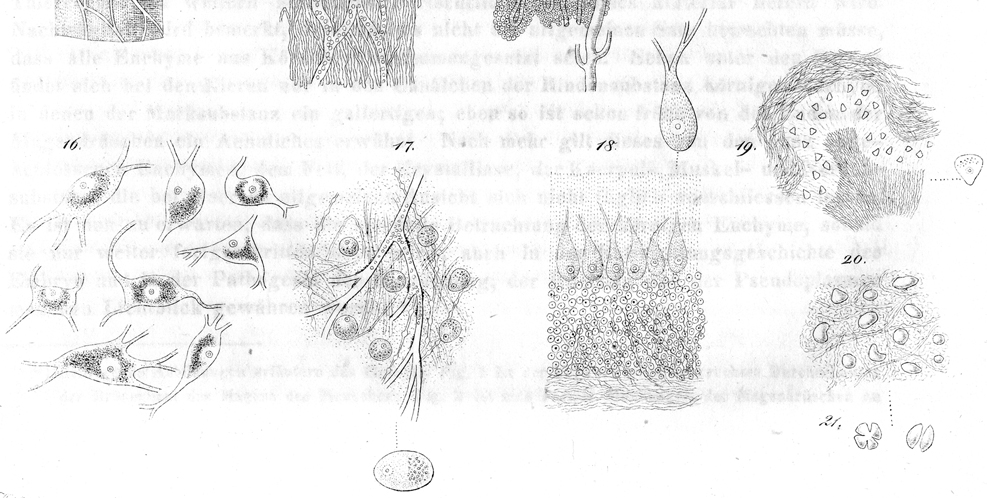
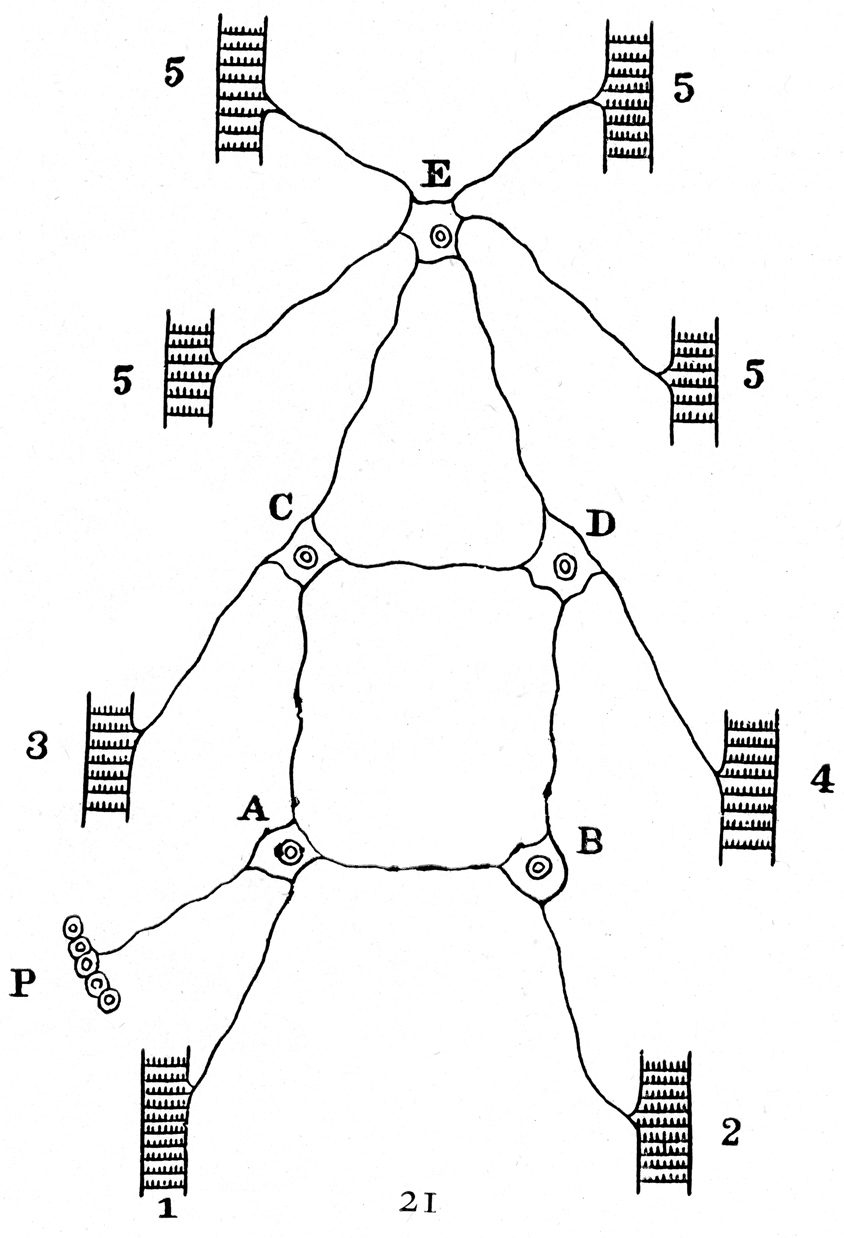
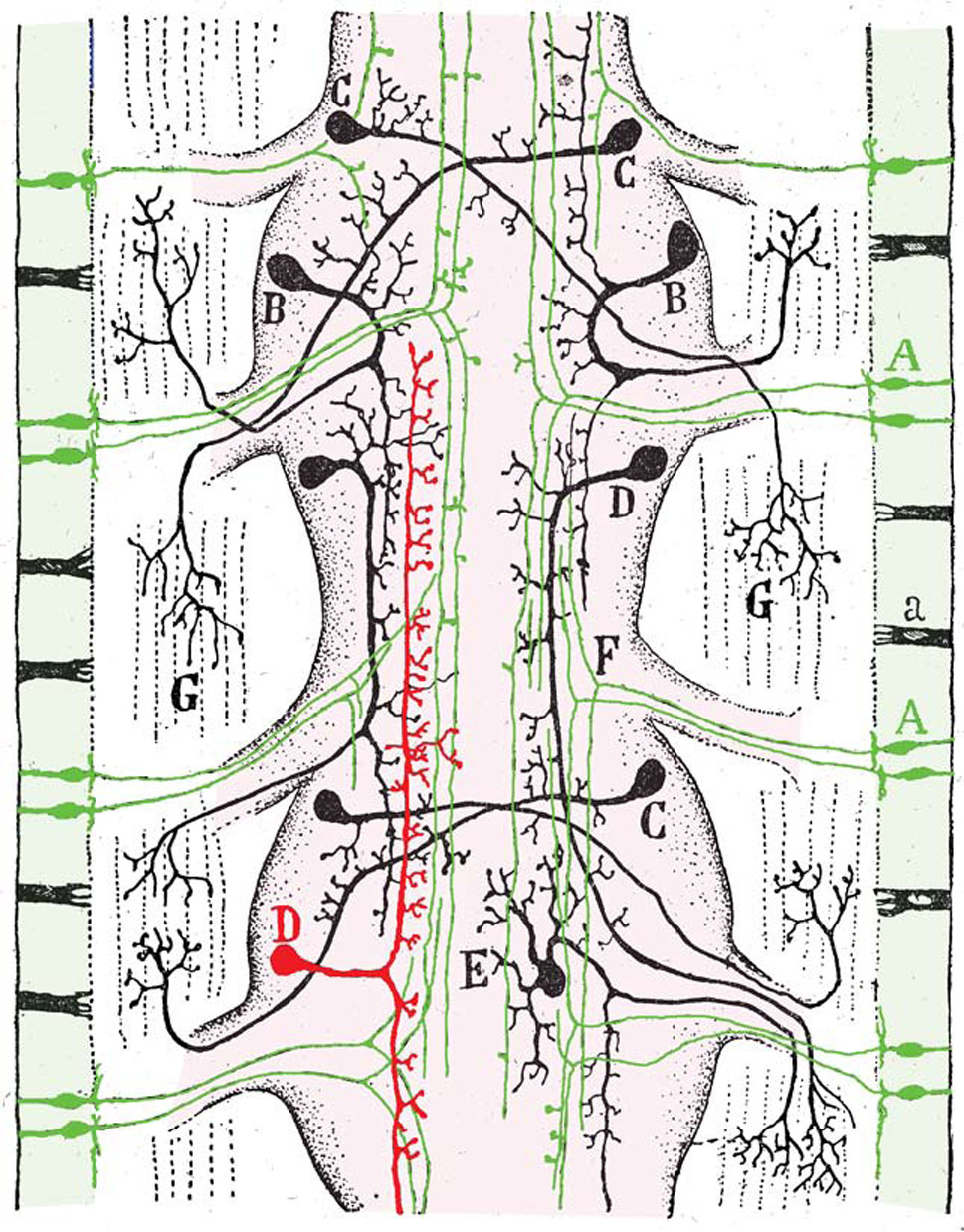
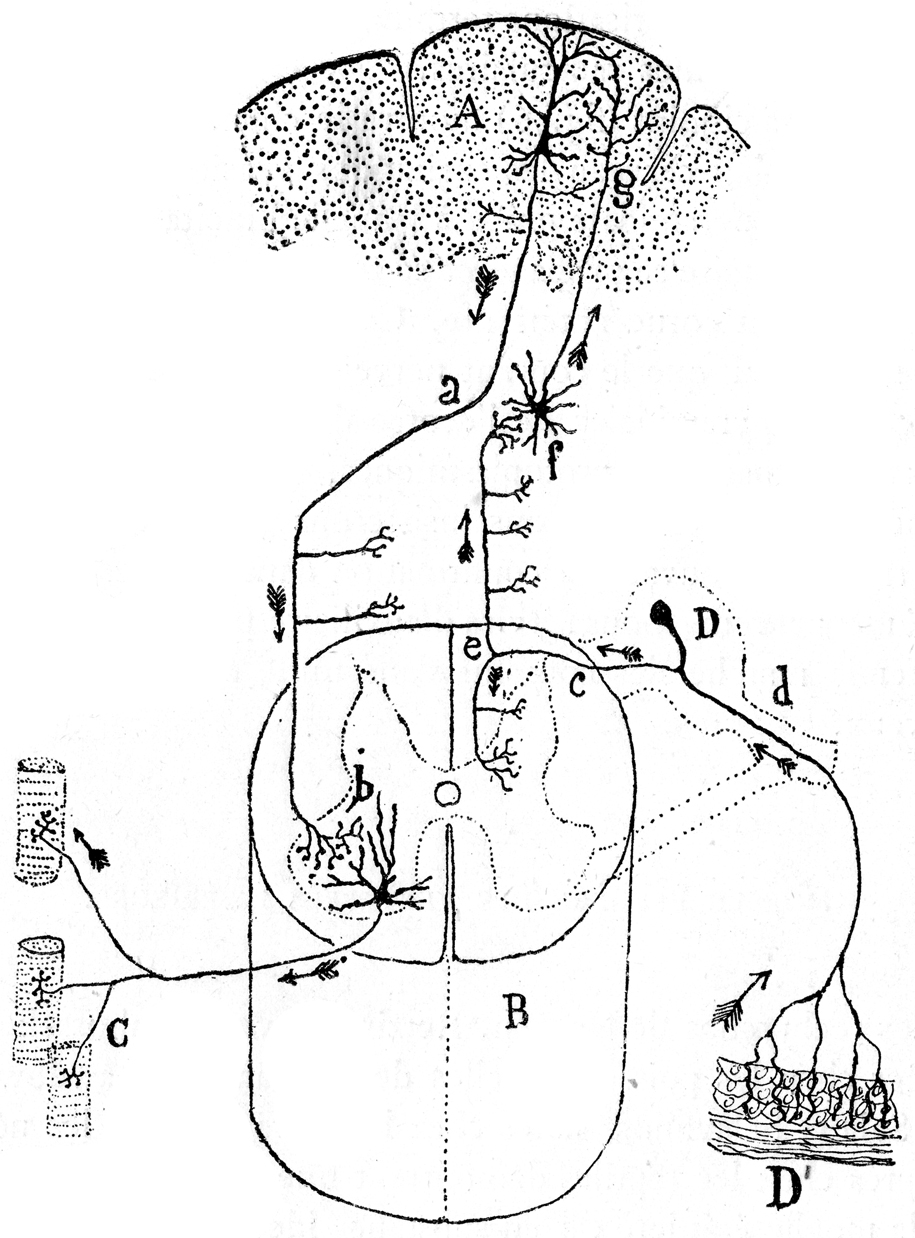
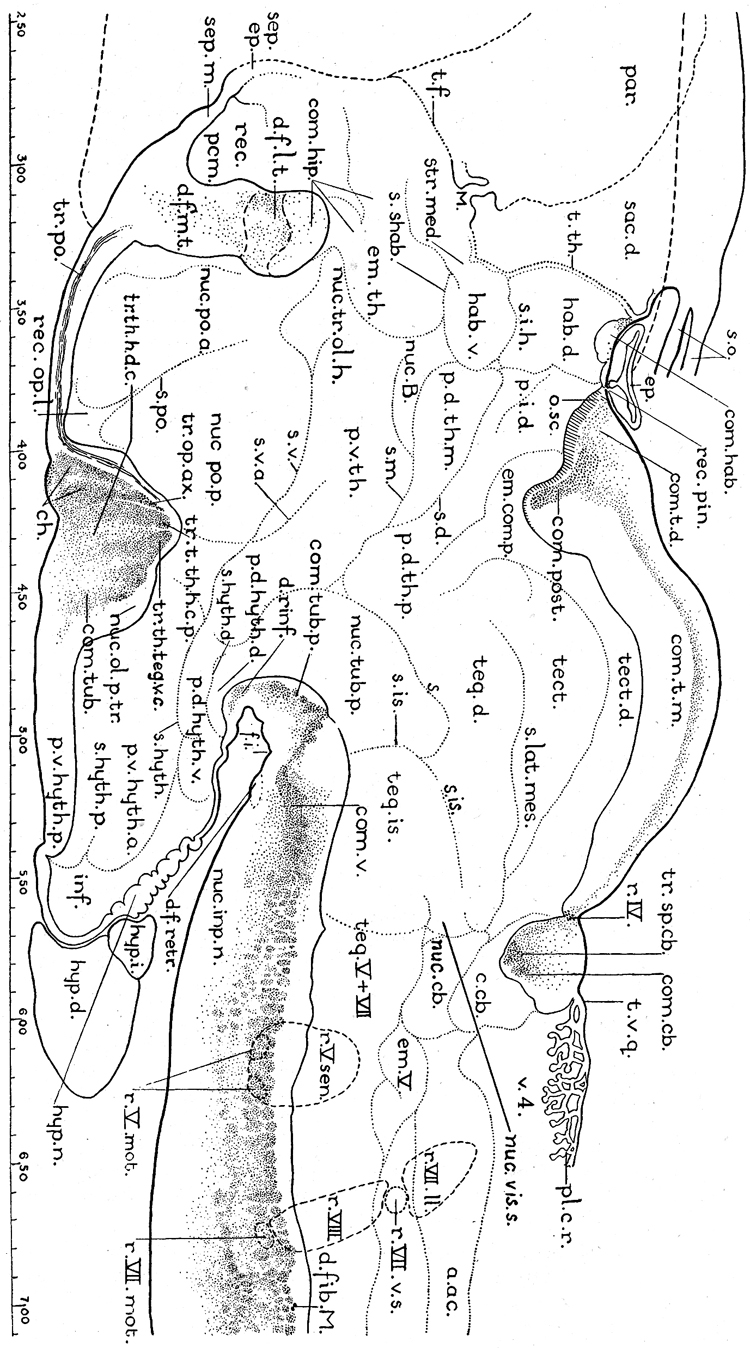
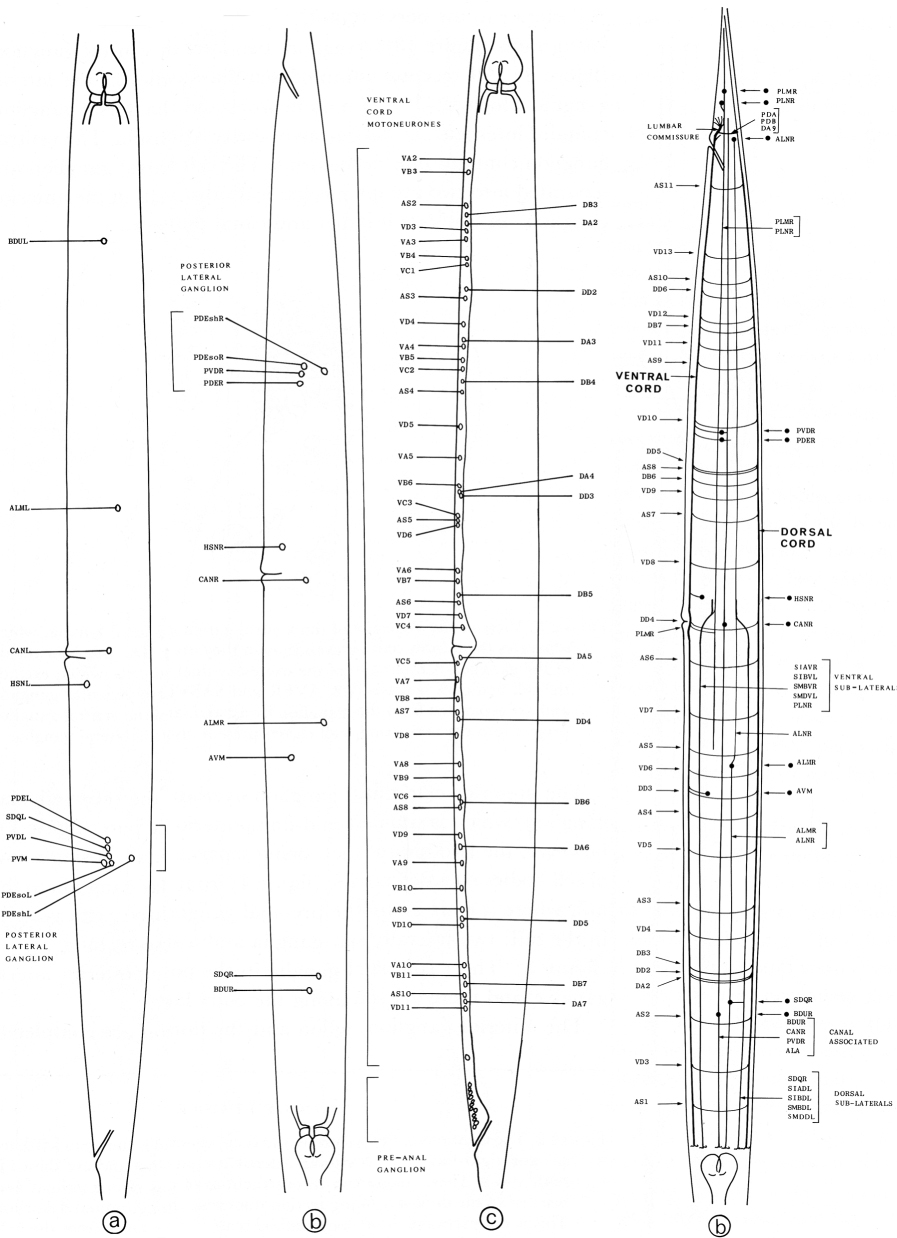
References
-
- Albertus Magnus. Philosophia pauperum sive philosophia naturalis. Brescia: B. Farfengus; 1490.
-
- Alvarez-Bolado G, Swanson LW. Developmental Brain Maps: Structure of the Embryonic Rat Brain. Amsterdam: Elsevier; 1996.
-
- Arbib MA, Grethe JG, editors. Computing the Brain: A Guide to Neuroinformatics. San Diego: Academic Press; 2001.
-
- Björklund A, Hökfelt T, editors. Handbook of Chemical Neuroanatomy. Amsterdam: Elsevier; 1983–2005.
-
- Bota M, Dong H-W, Swanson LW. From gene networks to brain networks. Nature Neurosci. 2003;6:795–799. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
