

RNA chaperone activity and RNA-binding properties of the E. coli protein StpA
- PMID: 17267410
- PMCID: PMC1851640
- DOI: 10.1093/nar/gkl1143
RNA chaperone activity and RNA-binding properties of the E. coli protein StpA
Abstract
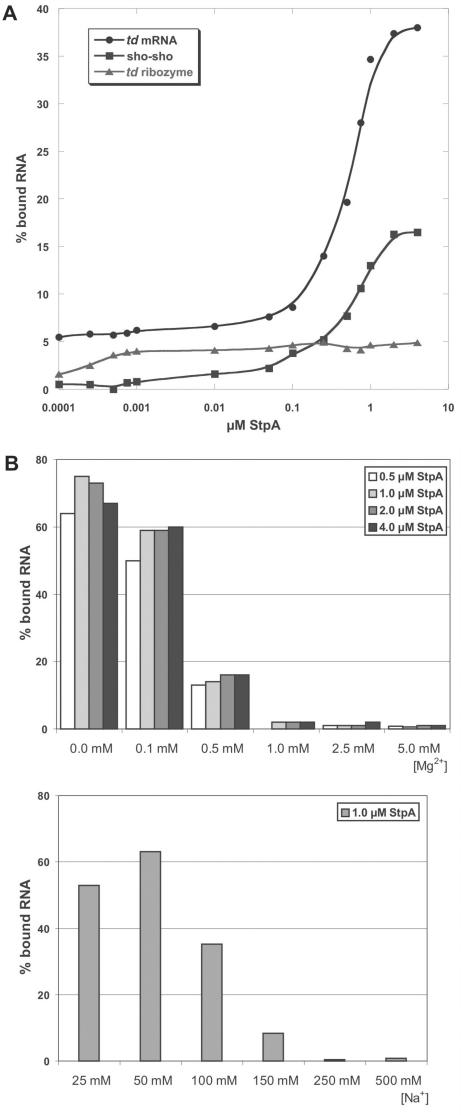
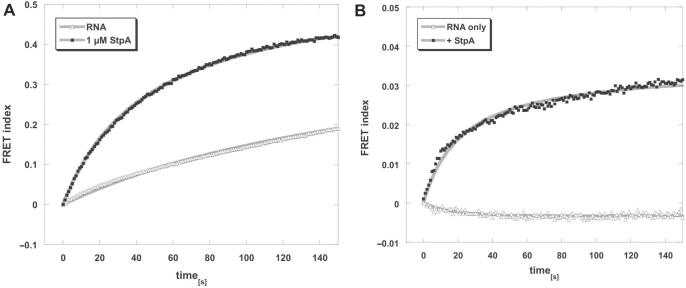
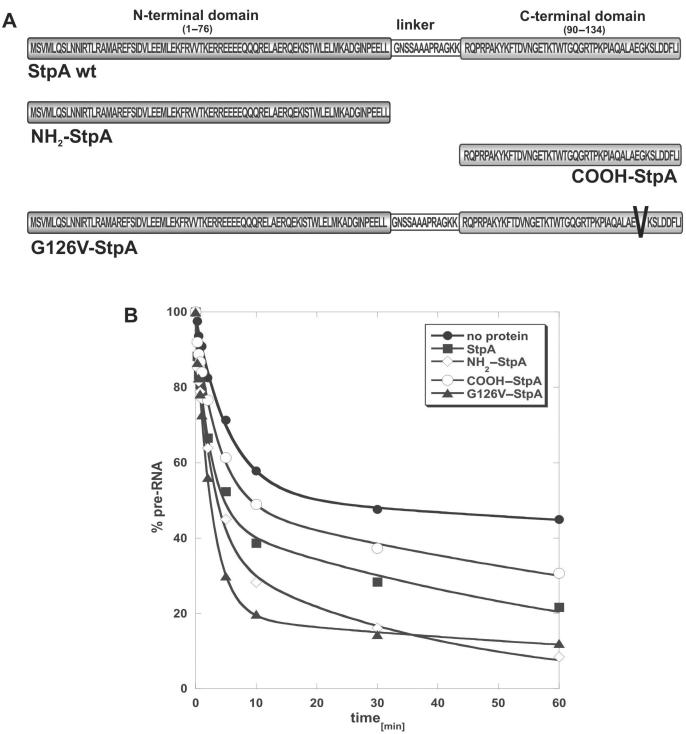
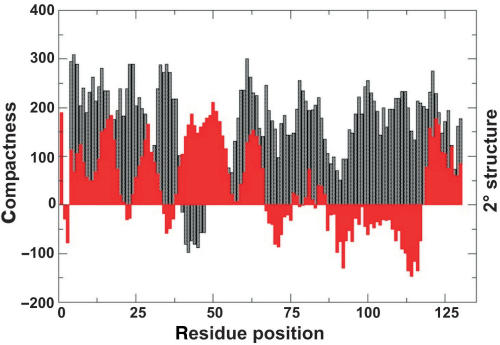
The E. coli protein StpA has RNA annealing and strand displacement activities and it promotes folding of RNAs by loosening their structures. To understand the mode of action of StpA, we analysed the relationship of its RNA chaperone activity to its RNA-binding properties. For acceleration of annealing of two short RNAs, StpA binds both molecules simultaneously, showing that annealing is promoted by crowding. StpA binds weakly to RNA with a preference for unstructured molecules. Binding of StpA to RNA is strongly dependent on the ionic strength, suggesting that the interactions are mainly electrostatic. A mutant variant of the protein, with a glycine to valine change in the nucleic-acid-binding domain, displays weaker RNA binding but higher RNA chaperone activity. This suggests that the RNA chaperone activity of StpA results from weak and transient interactions rather than from tight binding to RNA. We further discuss the role that structural disorder in proteins may play in chaperoning RNA folding, using bioinformatic sequence analysis tools, and provide evidence for the importance of conformational disorder and local structural preformation of chaperone nucleic-acid-binding sites.
Figures




References
-
- Weeks KM. Protein-facilitated RNA folding. Curr. Opin. Struct. Biol. 1997;7:336–342. - PubMed
-
- Schroeder R, Barta A, Semrad K. Strategies for RNA folding and assembly. Nat. Rev. Mol. Cell. Biol. 2004;5:908–919. - PubMed
-
- Coetzee T, Herschlag D, Belfort M. Escherichia coli proteins, including ribosomal protein S12, facilitate in vitro splicing of phage T4 introns by acting as RNA chaperones. Genes Dev. 1994;8:1575–1588. - PubMed
-
- Herschlag D. RNA chaperones and the RNA folding problem. J. Biol. Chem. 1995;270:20871–20874. - PubMed
-
- Cristofari G, Darlix JL. The ubiquitous nature of RNA chaperone proteins. Prog. Nucleic Acid Res. Mol. Biol. 2002;72:223–268. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
