

Herpesvirus saimiri episomal persistence is maintained via interaction between open reading frame 73 and the cellular chromosome-associated protein MeCP2
- PMID: 17267510
- PMCID: PMC1866103
- DOI: 10.1128/JVI.02171-06
Herpesvirus saimiri episomal persistence is maintained via interaction between open reading frame 73 and the cellular chromosome-associated protein MeCP2
Abstract
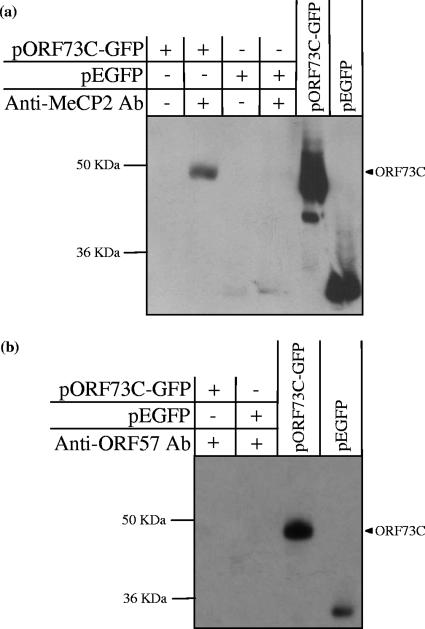
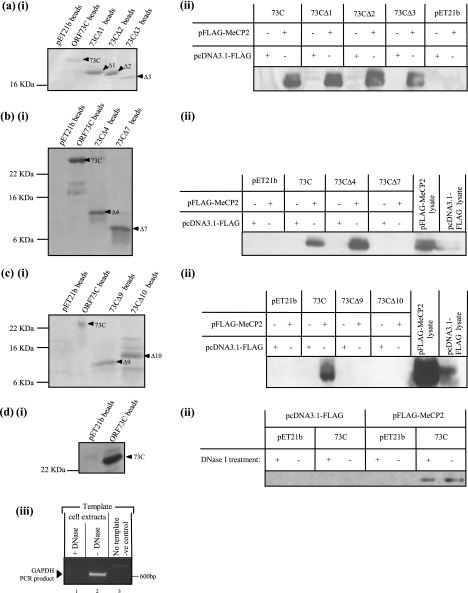
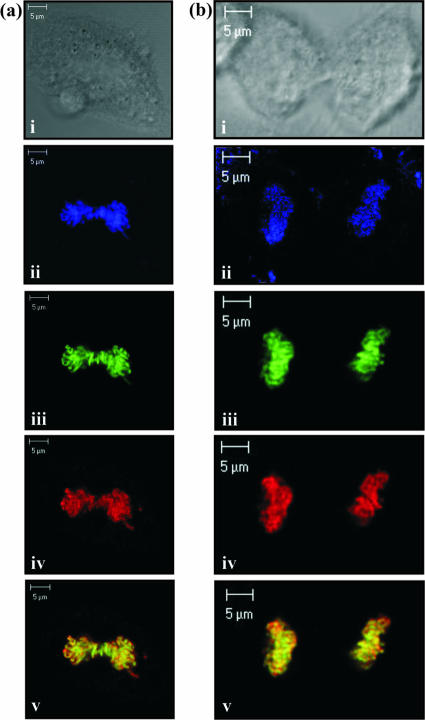
Herpesvirus saimiri (HVS) is the prototype gamma-2 herpesvirus, which naturally infects the squirrel monkey Saimiri sciureus, causing an asymptomatic but persistent infection. The latent phase of gamma-2 herpesviruses is characterized by their ability to persist in a dividing cell population while expressing a limited subset of latency-associated genes. In HVS only three genes, open reading frame 71 (ORF71), ORF72, and ORF73, are expressed from a polycistronic mRNA. ORF73 has been shown to be the only gene essential for HVS episomal maintenance and can therefore be functionally compared to the human gammaherpesvirus latency-associated proteins, EBNA-1 and Kaposi's sarcoma-associated herpesvirus (KSHV) latency-associated nuclear antigen (LANA). HVS ORF73 is the positional homologue of KSHV LANA and, although it shares limited sequence homology, has significant structural and functional similarities. Investigation of KSHV LANA has demonstrated that it is able to mediate KSHV episomal persistence by tethering the KSHV episome to host mitotic chromosomes via interactions with cellular chromosome-associated proteins. These include associations with core and linker histones, several bromodomain proteins, and the chromosome-associated proteins methyl CpG binding protein 2 (MeCP2) and DEK. Here we show that HVS ORF73 associates with MeCP2 via a 72-amino-acid domain within the ORF73 C terminus. Furthermore, we have assessed the functional significance of this interaction, using a variety of techniques including small hairpin RNA knockdown, and show that association between ORF73 and MeCP2 is essential for HVS chromosomal attachment and episomal persistence.
Figures




Similar articles
-
ORF73 of herpesvirus Saimiri strain C488 tethers the viral genome to metaphase chromosomes and binds to cis-acting DNA sequences in the terminal repeats.J Virol. 2003 Dec;77(23):12494-506. doi: 10.1128/jvi.77.23.12494-12506.2003. J Virol. 2003. PMID: 14610173 Free PMC article.
-
Accumulation of heterochromatin components on the terminal repeat sequence of Kaposi's sarcoma-associated herpesvirus mediated by the latency-associated nuclear antigen.J Virol. 2004 Jul;78(14):7299-310. doi: 10.1128/JVI.78.14.7299-7310.2004. J Virol. 2004. PMID: 15220403 Free PMC article.
-
Open reading frame 73 is required for herpesvirus saimiri A11-S4 episomal persistence.J Gen Virol. 2005 Oct;86(Pt 10):2703-2708. doi: 10.1099/vir.0.81230-0. J Gen Virol. 2005. PMID: 16186223
-
Herpesvirus saimiri.Philos Trans R Soc Lond B Biol Sci. 2001 Apr 29;356(1408):545-67. doi: 10.1098/rstb.2000.0780. Philos Trans R Soc Lond B Biol Sci. 2001. PMID: 11313011 Free PMC article. Review.
-
Genetic requirements for the episomal maintenance of oncogenic herpesvirus genomes.Adv Cancer Res. 2002;84:155-74. doi: 10.1016/s0065-230x(02)84005-2. Adv Cancer Res. 2002. PMID: 11883526 Review.
Cited by
-
The latency-associated nuclear antigen, a multifunctional protein central to Kaposi's sarcoma-associated herpesvirus latency.Future Microbiol. 2011 Dec;6(12):1399-413. doi: 10.2217/fmb.11.137. Future Microbiol. 2011. PMID: 22122438 Free PMC article. Review.
-
Potential of herpesvirus saimiri-based vectors to reprogram a somatic Ewing's sarcoma family tumor cell line.J Virol. 2013 Jun;87(12):7127-39. doi: 10.1128/JVI.03147-12. Epub 2013 Apr 17. J Virol. 2013. PMID: 23596304 Free PMC article.
-
The interaction of the gammaherpesvirus 68 orf73 protein with cellular BET proteins affects the activation of cell cycle promoters.J Virol. 2009 May;83(9):4423-34. doi: 10.1128/JVI.02274-08. Epub 2009 Feb 25. J Virol. 2009. PMID: 19244327 Free PMC article.
-
Merkel cell polyomavirus small T antigen targets the NEMO adaptor protein to disrupt inflammatory signaling.J Virol. 2013 Dec;87(24):13853-67. doi: 10.1128/JVI.02159-13. Epub 2013 Oct 9. J Virol. 2013. PMID: 24109239 Free PMC article.
-
The latency-associated nuclear antigen interacts with MeCP2 and nucleosomes through separate domains.J Virol. 2010 Mar;84(5):2318-30. doi: 10.1128/JVI.01097-09. Epub 2009 Dec 23. J Virol. 2010. PMID: 20032179 Free PMC article.
References
-
- Adkins, N. L., M. Watts, and P. T. Georgel. 2004. To the 30-nm chromatin fiber and beyond. Biochim. Biophys. Acta 1677:12-23. - PubMed
-
- Baer, R., A. T. Bankier, M. D. Biggin, P. L. Deininger, P. J. Farrell, T. J. Gibson, G. Hatfull, G. S. Hudson, S. C. Satchwell, C. Seguin, et al. 1984. DNA sequence and expression of the B95-8 Epstein-Barr virus genome. Nature 310:207-211. - PubMed
-
- Ballestas, M. E., P. A. Chatis, and K. M. Kaye. 1999. Efficient persistence of extrachromosomal KSHV DNA mediated by latency-associated nuclear antigen. Science 284:641-644. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
