

The nanovirus-encoded Clink protein affects plant cell cycle regulation through interaction with the retinoblastoma-related protein
- PMID: 17267511
- PMCID: PMC1866090
- DOI: 10.1128/JVI.02103-06
The nanovirus-encoded Clink protein affects plant cell cycle regulation through interaction with the retinoblastoma-related protein
Abstract
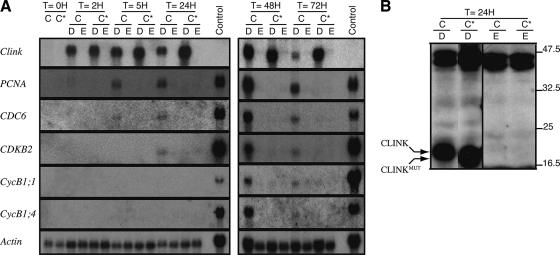
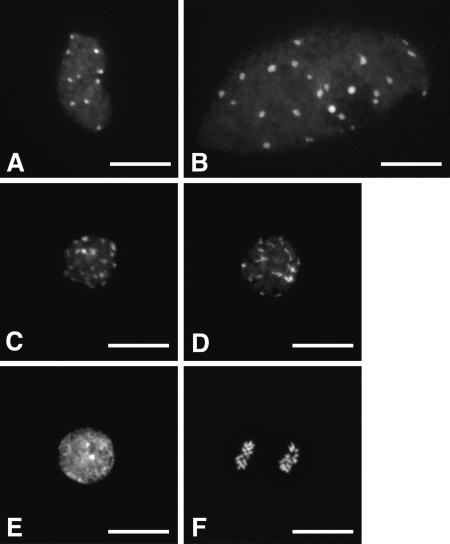
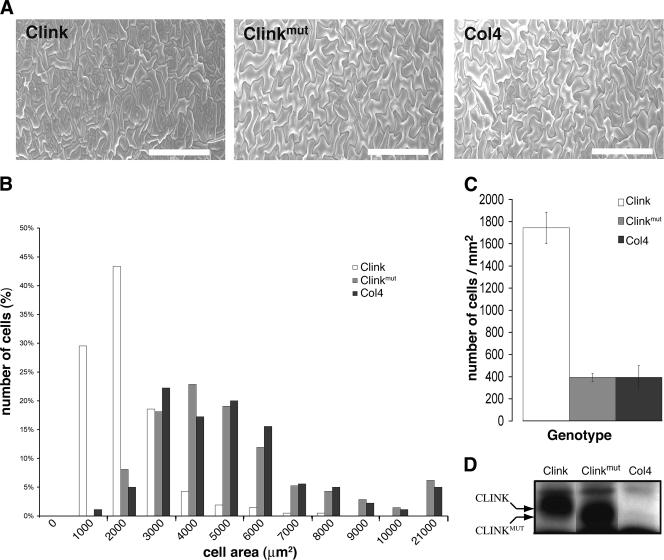
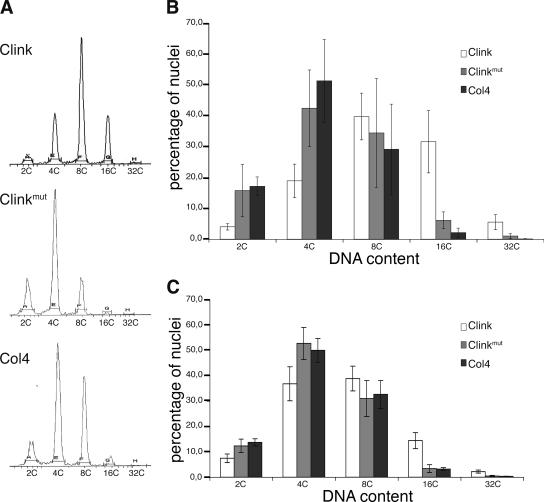
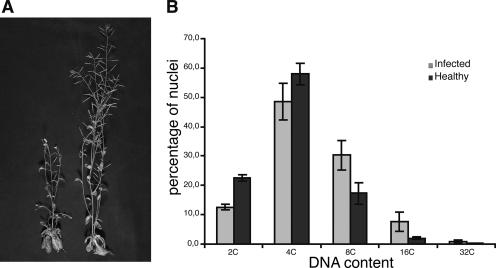
Nanoviruses, multicomponent single-stranded DNA plant viruses, encode a unique cell cycle link protein, Clink, that interacts with retinoblastoma-related proteins (RBR). We have established transgenic Arabidopsis thaliana lines that conditionally express Clink or a Clink variant deficient in RBR binding. By controlled induction of Clink expression, we demonstrated the capacity of the Clink protein to alter RBR function in vivo. We showed that transcription of both S-phase-specific and G2/M-phase-specific genes was up-regulated depending on the RBR-binding proficiency of Clink. Concomitantly, ploidy levels increased in a substantial fraction of leaf cell nuclei. Also, leaf epidermis cells of transgenic plants producing Clink were smaller and more numerous, indicating additional cell divisions in this tissue. Furthermore, cytogenetic analyses following induction of Clink expression in mature leaves revealed the presence of metaphasic and anaphasic nuclei, clear evidence that Clink-mediated RBR inactivation is sufficient to induce quiescent cells to reenter cell cycle progression and, for at least a fraction of them, to pass through mitosis. Expression of Clink had no effect on genes transcribed by RNA polymerases I and III, suggesting that, in contrast to its mammalian homologue, A. thaliana RBR is not involved in the repression of polymerase I and polymerase III transcription. The results of these in vivo analyses firmly establish Clink as a member of the diverse class of multifunctional cell cycle modulator proteins encoded by small DNA viruses.
Figures
Similar articles
-
Clink, a nanovirus-encoded protein, binds both pRB and SKP1.J Virol. 2000 Apr;74(7):2967-72. doi: 10.1128/jvi.74.7.2967-2972.2000. J Virol. 2000. PMID: 10708410 Free PMC article.
-
Cell type-specific role of the retinoblastoma/E2F pathway during Arabidopsis leaf development.Plant Physiol. 2006 Jan;140(1):67-80. doi: 10.1104/pp.105.071027. Epub 2005 Dec 16. Plant Physiol. 2006. PMID: 16361519 Free PMC article.
-
E2FB Interacts with RETINOBLASTOMA RELATED and Regulates Cell Proliferation during Leaf Development.Plant Physiol. 2020 Jan;182(1):518-533. doi: 10.1104/pp.19.00212. Epub 2019 Nov 6. Plant Physiol. 2020. PMID: 31694902 Free PMC article.
-
Nanoviruses: genome organisation and protein function.Vet Microbiol. 2004 Feb 4;98(2):103-9. doi: 10.1016/j.vetmic.2003.10.015. Vet Microbiol. 2004. PMID: 14741122 Review.
-
Arabidopsis RETINOBLASTOMA-RELATED and Polycomb group proteins: cooperation during plant cell differentiation and development.J Exp Bot. 2014 Jun;65(10):2667-76. doi: 10.1093/jxb/eru069. Epub 2014 Mar 17. J Exp Bot. 2014. PMID: 24638900 Review.
Cited by
-
The ubiquitin/26S proteasome system in plant-pathogen interactions: a never-ending hide-and-seek game.Mol Plant Pathol. 2010 Mar;11(2):293-308. doi: 10.1111/j.1364-3703.2009.00596.x. Mol Plant Pathol. 2010. PMID: 20447278 Free PMC article. Review.
-
Geminiviruses subvert ubiquitination by altering CSN-mediated derubylation of SCF E3 ligase complexes and inhibit jasmonate signaling in Arabidopsis thaliana.Plant Cell. 2011 Mar;23(3):1014-32. doi: 10.1105/tpc.110.080267. Epub 2011 Mar 25. Plant Cell. 2011. PMID: 21441437 Free PMC article.
-
Reassortments in single-stranded DNA multipartite viruses: Confronting expectations based on molecular constraints with field observations.Virus Evol. 2024 Feb 2;10(1):veae010. doi: 10.1093/ve/veae010. eCollection 2024. Virus Evol. 2024. PMID: 38384786 Free PMC article. Review.
-
Plant geminivirus rep protein induces rereplication in fission yeast.J Virol. 2009 Jul;83(13):6769-78. doi: 10.1128/JVI.02491-08. Epub 2009 Apr 15. J Virol. 2009. PMID: 19369323 Free PMC article.
-
Interaction of CPR5 with cell cycle regulators UVI4 and OSD1 in Arabidopsis.PLoS One. 2014 Jun 19;9(6):e100347. doi: 10.1371/journal.pone.0100347. eCollection 2014. PLoS One. 2014. PMID: 24945150 Free PMC article.
References
-
- Accotto, G. P., P. M. Mullineaux, S. C. Brown, and D. Marie. 1993. Digitaria streak geminivirus replicative forms are abundant in S-phase nuclei of infected cells. Virology 195:257-259. - PubMed
-
- Aoyama, T., and N. H. Chua. 1997. A glucocorticoid-mediated transcriptional induction system in transgenic plants. Plant J. 11:605-612. - PubMed
-
- Aronson, M. 2002. Ph.D. thesis. Institut National d'Agronomie Paris-Grignon, Paris, France.
-
- Aronson, M. N., A. Complainville, D. Clerot, H. Alcalde, L. Katul, H. J. Vetten, B. Gronenborn, and T. Timchenko. 2002. In planta protein-protein interactions assessed using a nanovirus-based replication and expression system. Plant J. 31:767-775. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
