

Sensory-motor transformation by individual command neurons
- PMID: 17267556
- PMCID: PMC6673182
- DOI: 10.1523/JNEUROSCI.4925-06.2007
Sensory-motor transformation by individual command neurons
Abstract
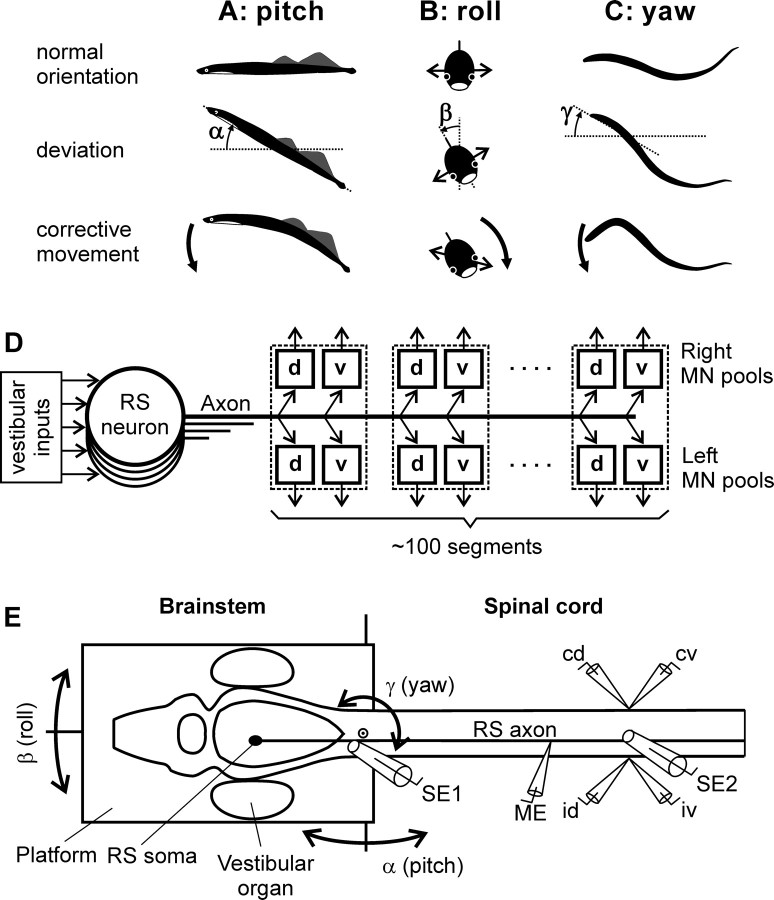
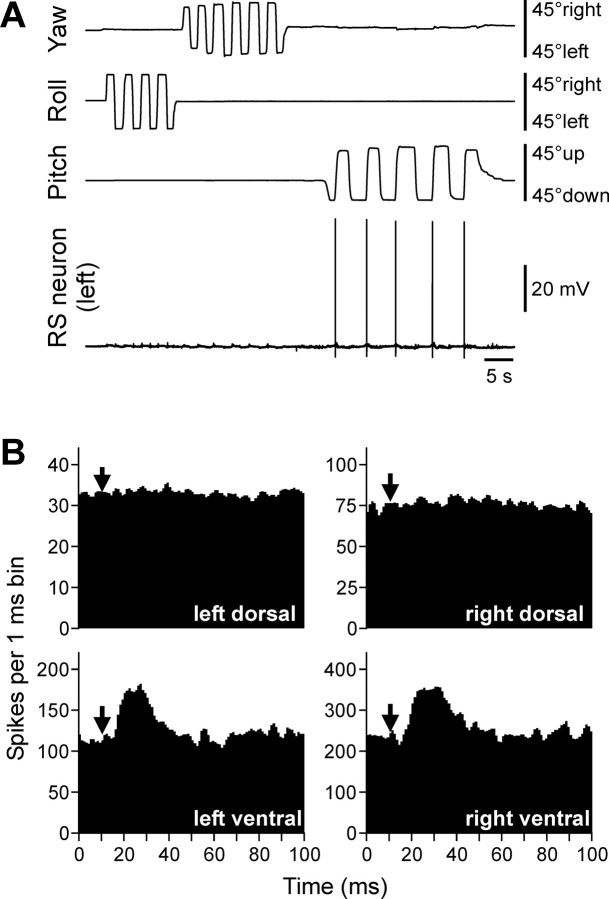
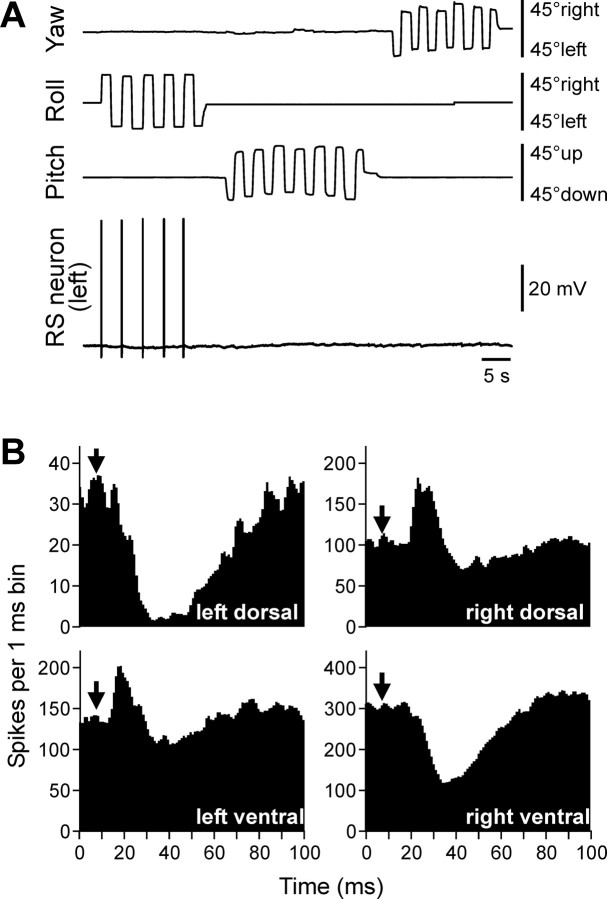
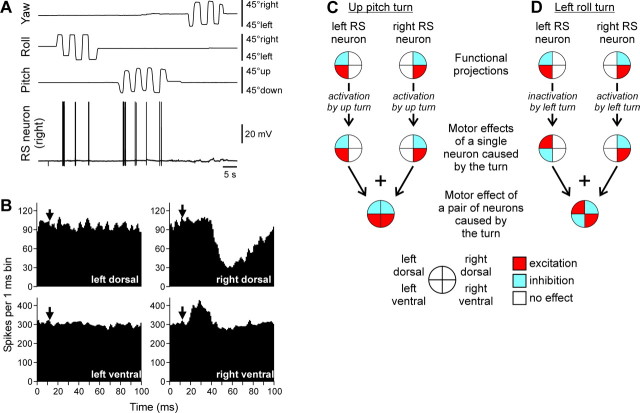
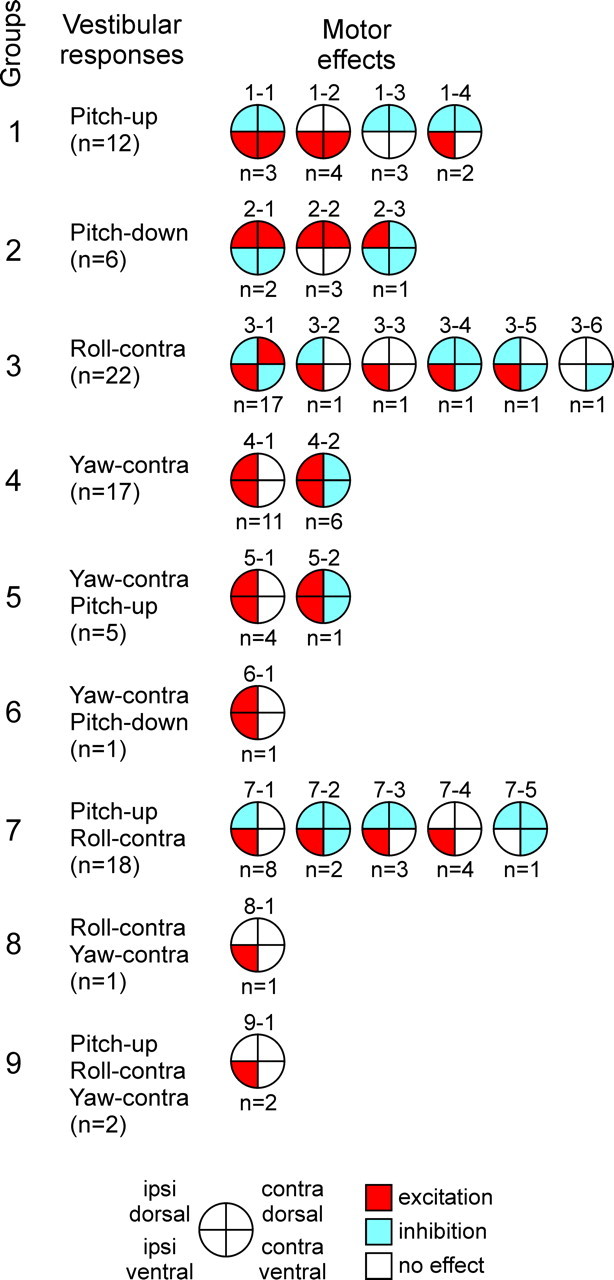
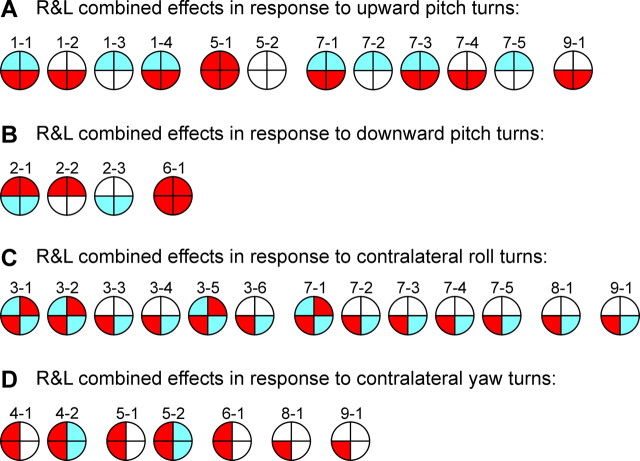
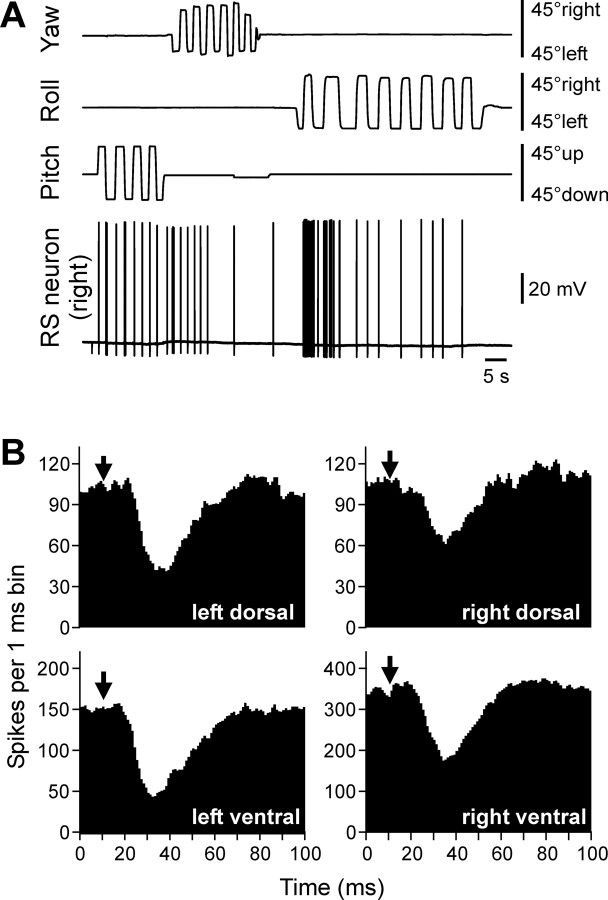
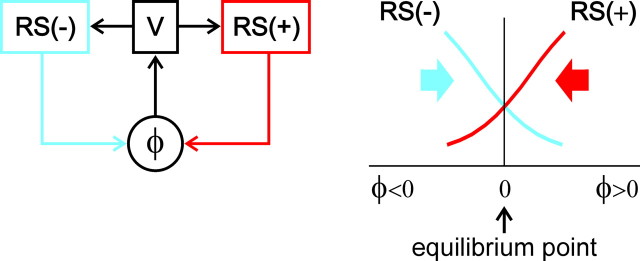
Animals and humans maintain a definite body orientation in space during locomotion. Here we analyze the system for the control of body orientation in the lamprey (a lower vertebrate). In the swimming lamprey, commands for changing the body orientation are based on vestibular information; they are transmitted to the spinal cord by reticulospinal (RS) neurons. The aim of this study was to characterize the sensory-motor transformation performed by individual RS neurons. The brainstem-spinal cord preparation with vestibular organs was used. For each RS neuron, we recorded (1) its vestibular responses to turns in different planes and (2) responses in different motoneuron pools of the spinal cord to stimulation of the same RS neuron; the latter data allowed us to estimate the direction of torque (caused by the RS neuron) that will rotate the animal's body during swimming. For each of the three main planes (roll, pitch, and yaw), two groups of RS neurons were found; they were activated by rotation in opposite directions and caused the torques counteracting the rotation that activated the neuron. In each plane, the system will stabilize the orientation at which the two groups are equally active; any deviation from this orientation will evoke a corrective motor response. Thus, individual RS neurons transform sensory information about the body orientation into the motor commands that cause corrections of orientation. The closed-loop mechanisms formed by individual neurons of a group operate in parallel to generate the resulting motor responses.
Figures
Similar articles
-
Responses of reticulospinal neurons in intact lamprey to pitch tilt.J Neurophysiol. 2002 Sep;88(3):1136-46. doi: 10.1152/jn.2002.88.3.1136. J Neurophysiol. 2002. PMID: 12205135
-
Postural control in the lamprey: A study with a neuro-mechanical model.J Neurophysiol. 2000 Dec;84(6):2880-7. doi: 10.1152/jn.2000.84.6.2880. J Neurophysiol. 2000. PMID: 11110817
-
Responses of reticulospinal neurons in intact lamprey to vestibular and visual inputs.J Neurophysiol. 2000 Feb;83(2):864-78. doi: 10.1152/jn.2000.83.2.864. J Neurophysiol. 2000. PMID: 10669500
-
Spinal and supraspinal postural networks.Brain Res Rev. 2008 Jan;57(1):212-21. doi: 10.1016/j.brainresrev.2007.06.017. Epub 2007 Jul 27. Brain Res Rev. 2008. PMID: 17822773 Free PMC article. Review.
-
Encoding and decoding of reticulospinal commands.Brain Res Brain Res Rev. 2002 Oct;40(1-3):166-77. doi: 10.1016/s0165-0173(02)00199-6. Brain Res Brain Res Rev. 2002. PMID: 12589915 Review.
Cited by
-
Putative spinal interneurons mediating postural limb reflexes provide a basis for postural control in different planes.Eur J Neurosci. 2015 Jan;41(2):168-81. doi: 10.1111/ejn.12780. Epub 2014 Nov 5. Eur J Neurosci. 2015. PMID: 25370349 Free PMC article.
-
The spinobulbar system in lamprey.Brain Res Rev. 2008 Jan;57(1):37-45. doi: 10.1016/j.brainresrev.2007.07.010. Epub 2007 Aug 6. Brain Res Rev. 2008. PMID: 17716741 Free PMC article. Review.
-
Physiological and circuit mechanisms of postural control.Curr Opin Neurobiol. 2012 Aug;22(4):646-52. doi: 10.1016/j.conb.2012.03.002. Epub 2012 Mar 23. Curr Opin Neurobiol. 2012. PMID: 22446009 Free PMC article. Review.
-
Brainstem neural mechanisms controlling locomotion with special reference to basal vertebrates.Front Neural Circuits. 2023 Mar 30;17:910207. doi: 10.3389/fncir.2023.910207. eCollection 2023. Front Neural Circuits. 2023. PMID: 37063386 Free PMC article. Review.
-
Role of CaMKIIa reticular neurons of caudal medulla in control of posture.bioRxiv [Preprint]. 2025 Mar 17:2025.03.17.643745. doi: 10.1101/2025.03.17.643745. bioRxiv. 2025. PMID: 40166225 Free PMC article. Preprint.
References
-
- Brodin L, Grillner S, Dubuc R, Ohta Y, Kasicki S, Hökfelt T. Reticulospinal neurons in lamprey: transmitters, synaptic interactions and their role during locomotion. Arch Ital Biol. 1988;126:317–345. - PubMed
-
- Bussières N. Etude Anatomique et Functionnelle. Montreal: University of Montreal; 1994. Les Systemes Descendants chez la Lamproie.
-
- Deliagina TG, Fagerstedt P. Responses of reticulospinal neurons in intact lamprey to vestibular and visual inputs. J Neurophysiol. 2000;83:864–878. - PubMed
-
- Deliagina TG, Orlovsky GN. Comparative neurobiology of postural control. Curr Opin Neurobiol. 2002;12:652–657. - PubMed
-
- Deliagina TG, Pavlova EL. Modifications of vestibular responses of individual reticulospinal neurons in the lamprey caused by a unilateral labyrinthectomy. J Neurophysiol. 2002;87:1–14. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources