

Neocortical inhibitory terminals innervate dendritic spines targeted by thalamocortical afferents
- PMID: 17267569
- PMCID: PMC6673192
- DOI: 10.1523/JNEUROSCI.3846-06.2007
Neocortical inhibitory terminals innervate dendritic spines targeted by thalamocortical afferents
Abstract
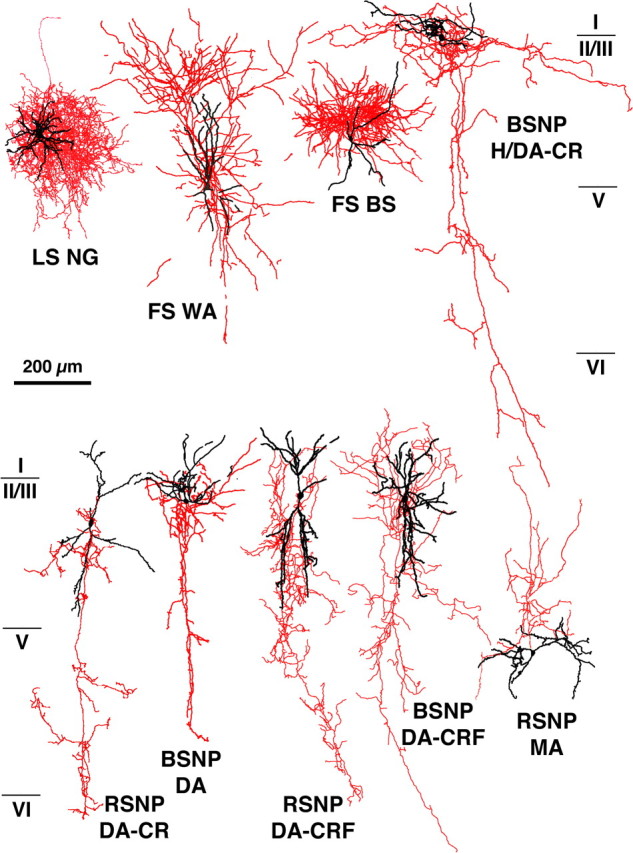
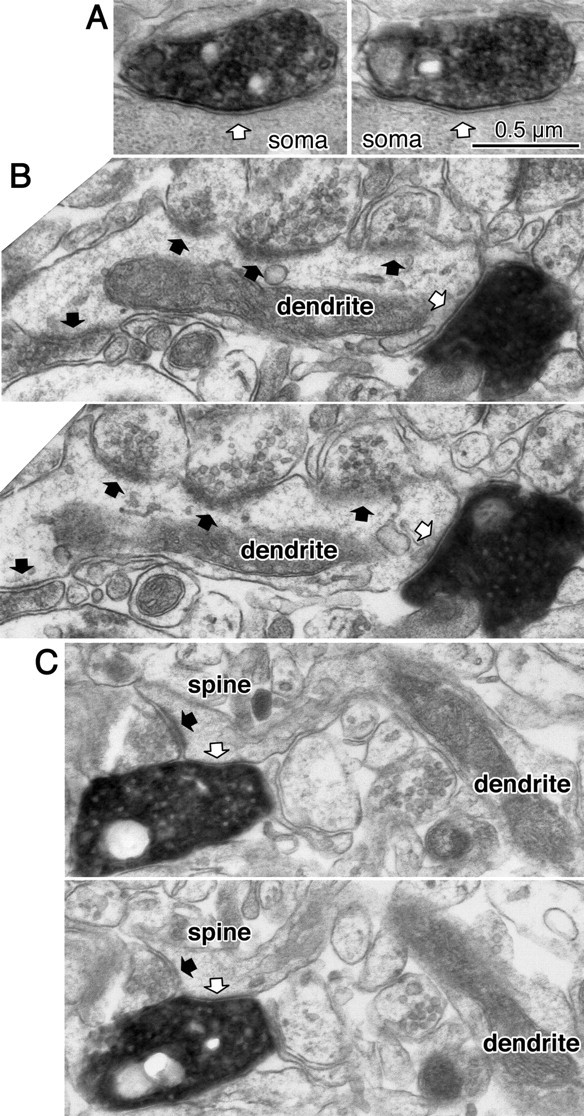
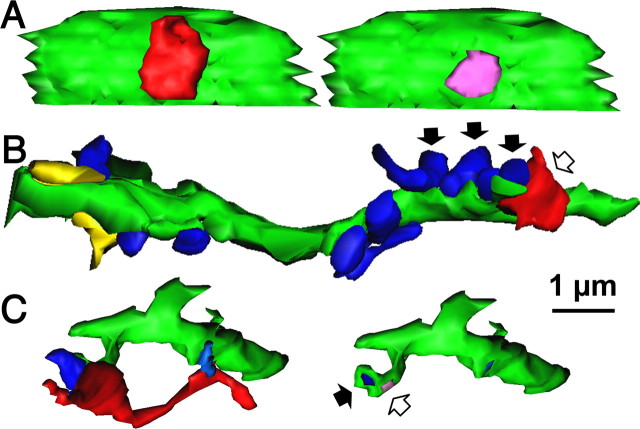
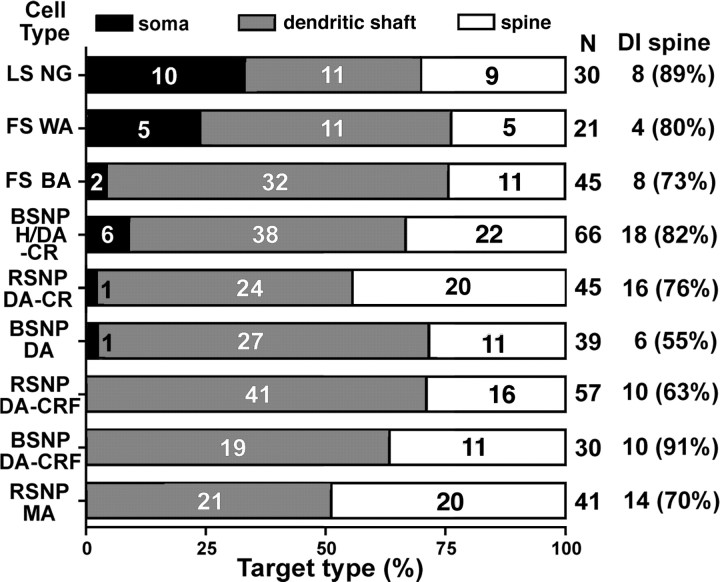
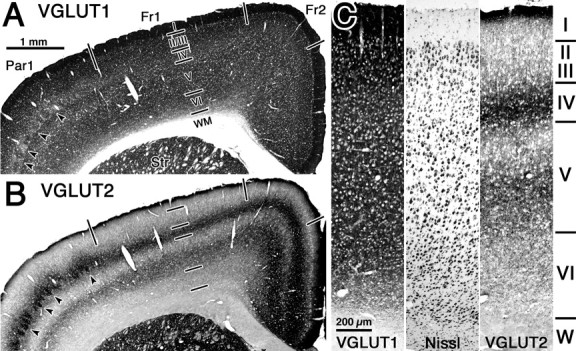
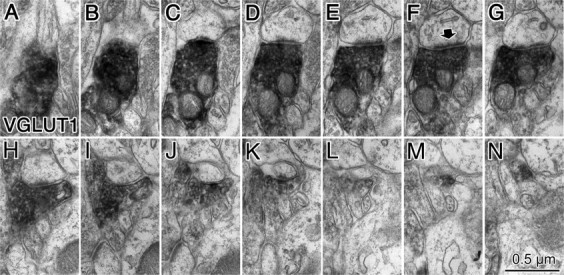
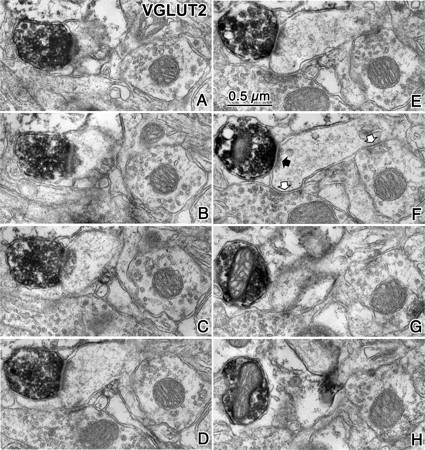
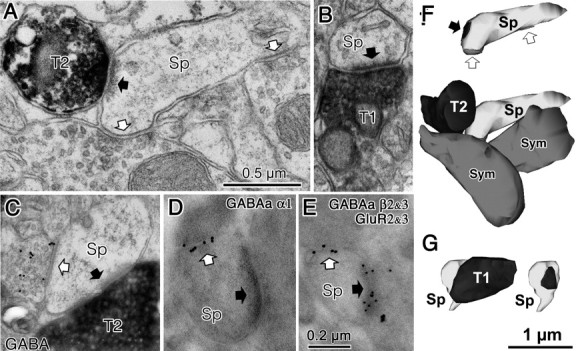
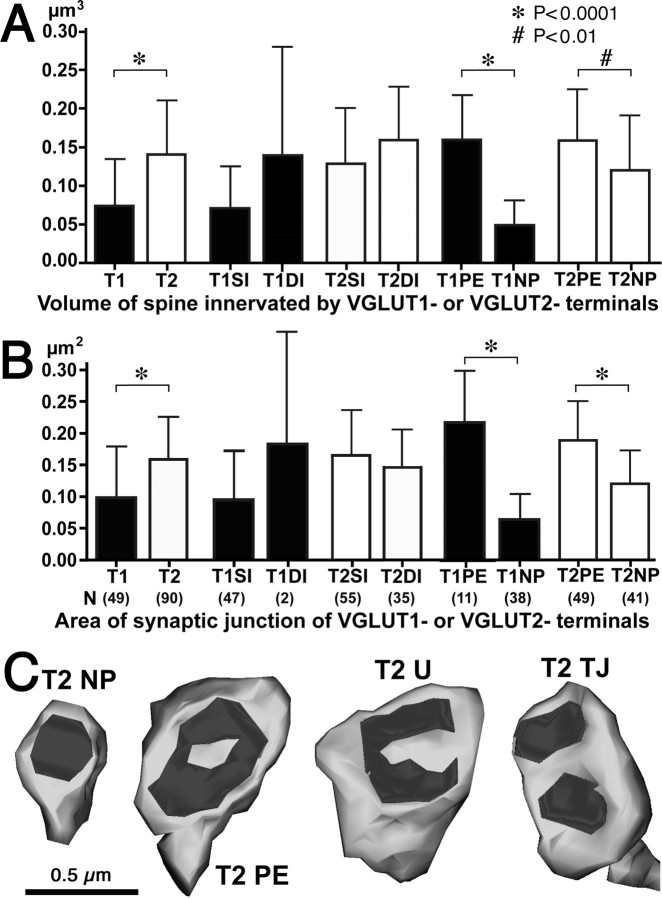
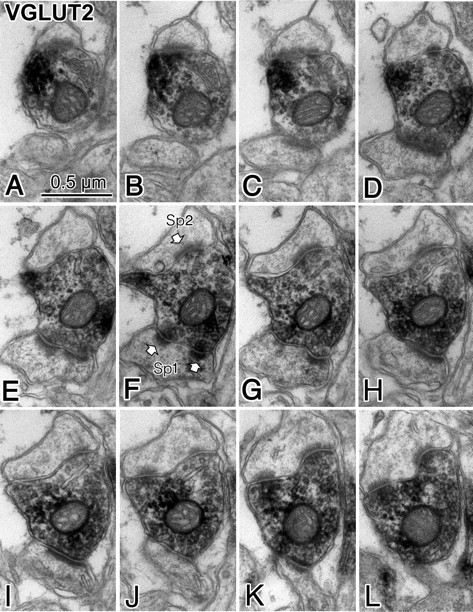
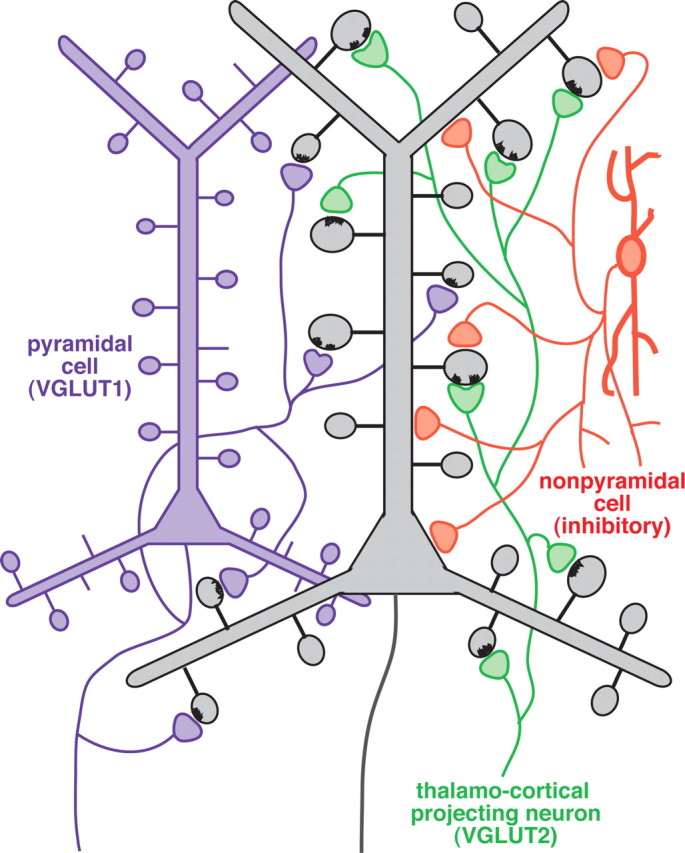
Fast inhibition in the cortex is gated primarily at GABAergic synapses formed by local interneurons onto postsynaptic targets. Although GABAergic inputs to the somata and axon initial segments of neocortical pyramidal neurons are associated with direct inhibition of action potential generation, the role of GABAergic inputs to distal dendritic segments, including spines, is less well characterized. Because a significant proportion of inhibitory input occurs on distal dendrites and spines, it will be important to determine whether these GABAergic synapses are formed selectively by certain classes of presynaptic cells onto specific postsynaptic elements. By electron microscopic observations of synapses formed by different subtypes of nonpyramidal cells, we found that a surprisingly large fraction (33.4 +/- 9.3%) of terminals formed symmetrical synaptic junctions onto a subset of cortical spines that were mostly coinnervated by an asymmetrical terminal. Using VGLUT1 and VGLUT2 isoform of the glutamate vesicular transporter immunohistochemistry, we found that the double-innervated spines selectively received thalamocortical afferents expressing the VGLUT2 but almost never intracortical inputs expressing the VGLUT1. When comparing the volumes of differentially innervated spines and their synaptic junction areas, we found that spines innervated by VGLUT2-positive terminal were significantly larger than spines innervated by VGLUT1-positive terminal and that these spines had larger, and more often perforated, synapses than those of spines innervated by VGLUT1-positive afferent. These results demonstrate that inhibitory inputs to pyramidal cell spines may preferentially reduce thalamocortical rather than intracortical synaptic transmission and are therefore positioned to selectively gate extracortical information.
Figures
Similar articles
-
Cortical and thalamic innervation of direct and indirect pathway medium-sized spiny neurons in mouse striatum.J Neurosci. 2010 Nov 3;30(44):14610-8. doi: 10.1523/JNEUROSCI.1623-10.2010. J Neurosci. 2010. PMID: 21048118 Free PMC article.
-
Fast IPSPs elicited via multiple synaptic release sites by different types of GABAergic neurone in the cat visual cortex.J Physiol. 1997 May 1;500 ( Pt 3)(Pt 3):715-38. doi: 10.1113/jphysiol.1997.sp022054. J Physiol. 1997. PMID: 9161987 Free PMC article.
-
Synaptic Targets and Cellular Sources of CB1 Cannabinoid Receptor and Vesicular Glutamate Transporter-3 Expressing Nerve Terminals in Relation to GABAergic Neurons in the Human Cerebral Cortex.Eur J Neurosci. 2025 Jan;61(1):e16652. doi: 10.1111/ejn.16652. Eur J Neurosci. 2025. PMID: 39810425 Free PMC article.
-
Connexon connexions in the thalamocortical system.Prog Brain Res. 2005;149:41-57. doi: 10.1016/S0079-6123(05)49004-4. Prog Brain Res. 2005. PMID: 16226575 Review.
-
Fast-spike interneurons and feedforward inhibition in awake sensory neocortex.Cereb Cortex. 2003 Jan;13(1):25-32. doi: 10.1093/cercor/13.1.25. Cereb Cortex. 2003. PMID: 12466212 Review.
Cited by
-
Control of synaptic plasticity in deep cortical networks.Nat Rev Neurosci. 2018 Feb 16;19(3):166-180. doi: 10.1038/nrn.2018.6. Nat Rev Neurosci. 2018. PMID: 29449713 Review.
-
Rapid, learning-induced inhibitory synaptogenesis in murine barrel field.J Neurosci. 2010 Jan 20;30(3):1176-84. doi: 10.1523/JNEUROSCI.2970-09.2010. J Neurosci. 2010. PMID: 20089926 Free PMC article.
-
The effects of aging on neuropil structure in mouse somatosensory cortex-A 3D electron microscopy analysis of layer 1.PLoS One. 2018 Jul 2;13(7):e0198131. doi: 10.1371/journal.pone.0198131. eCollection 2018. PLoS One. 2018. PMID: 29966021 Free PMC article.
-
SRGAP2 and Its Human-Specific Paralog Co-Regulate the Development of Excitatory and Inhibitory Synapses.Neuron. 2016 Jul 20;91(2):356-69. doi: 10.1016/j.neuron.2016.06.013. Epub 2016 Jun 30. Neuron. 2016. PMID: 27373832 Free PMC article.
-
Reduced cortical innervation of the subthalamic nucleus in MPTP-treated parkinsonian monkeys.Brain. 2015 Apr;138(Pt 4):946-62. doi: 10.1093/brain/awv018. Epub 2015 Feb 12. Brain. 2015. PMID: 25681412 Free PMC article.
References
-
- Beaulieu C, Kisvárday Z, Somogyi P, Cynader M, Cowey A. Quantitative distribution of GABA-immunopositive and -immunonegative neurons and synapses in the monkey striate cortex (area 17) Cereb Cortex. 1992;2:295–309. - PubMed
-
- Berendse HW, Groenewegen HJ. Restricted cortical termination fields of the midline and intralaminar thalamic nuclei in the rat. Neuroscience. 1991;42:73–102. - PubMed
-
- Bruno RM, Sakmann B. Cortex is driven by weak but synchronously active thalamocortical synapses. Science. 2006;312:1622–1627. - PubMed
-
- Buhl EH, Halasy K, Somogyi P. Diverse sources of hippocampal unitary inhibitory postsynaptic potentials and the number of synaptic release sites. Nature. 1994;368:823–828. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources