

Reconstitution and subunit geometry of human condensin complexes
- PMID: 17268547
- PMCID: PMC1852836
- DOI: 10.1038/sj.emboj.7601562
Reconstitution and subunit geometry of human condensin complexes
Abstract
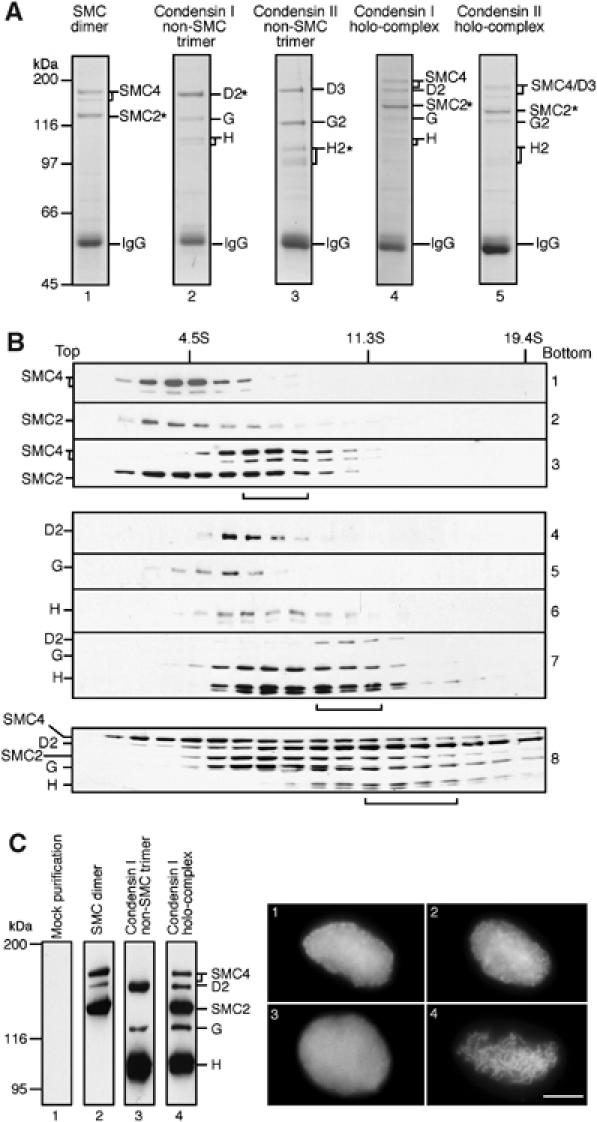
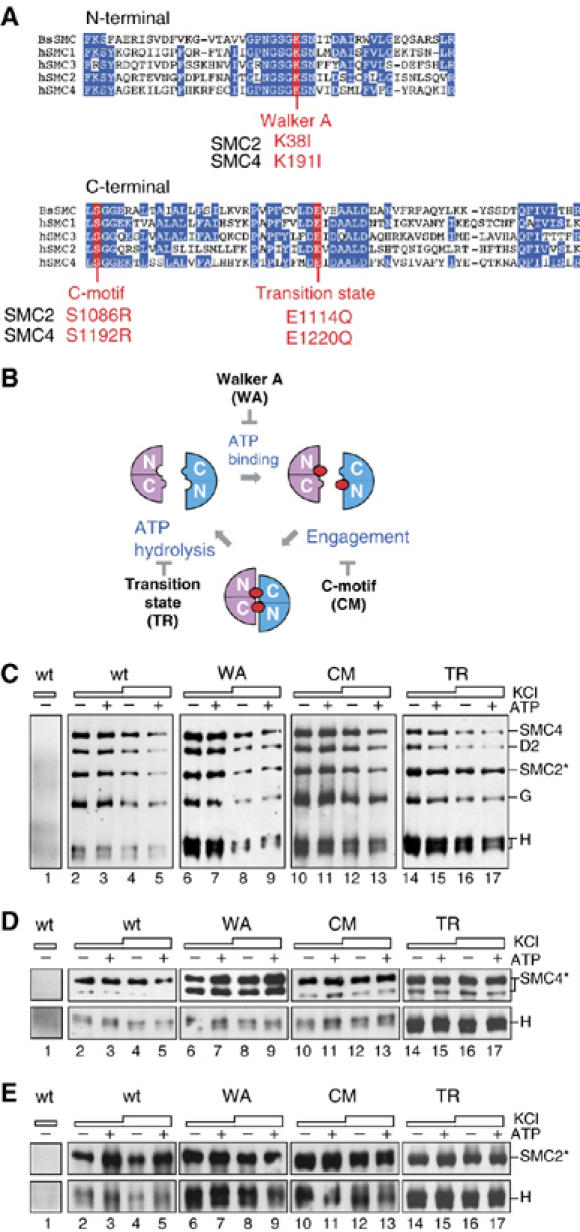
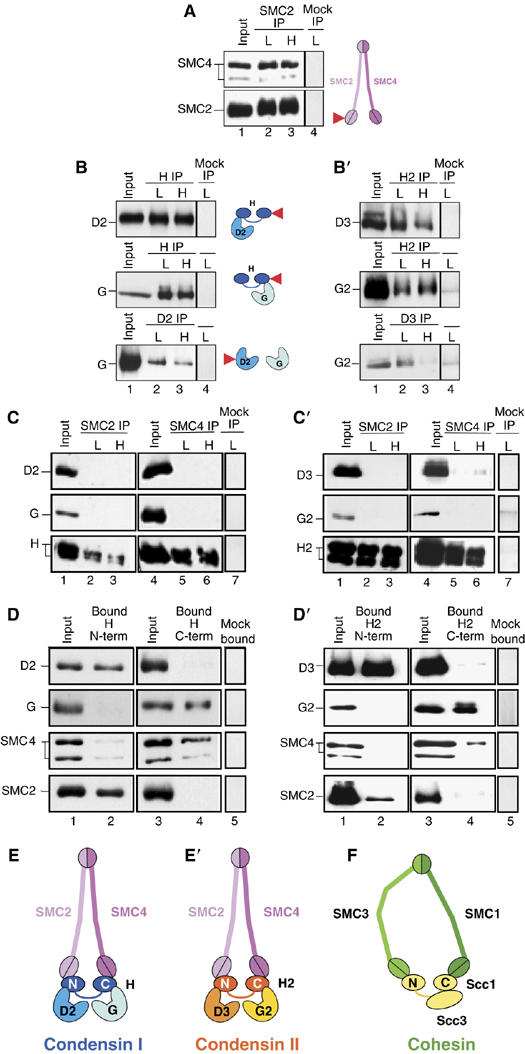
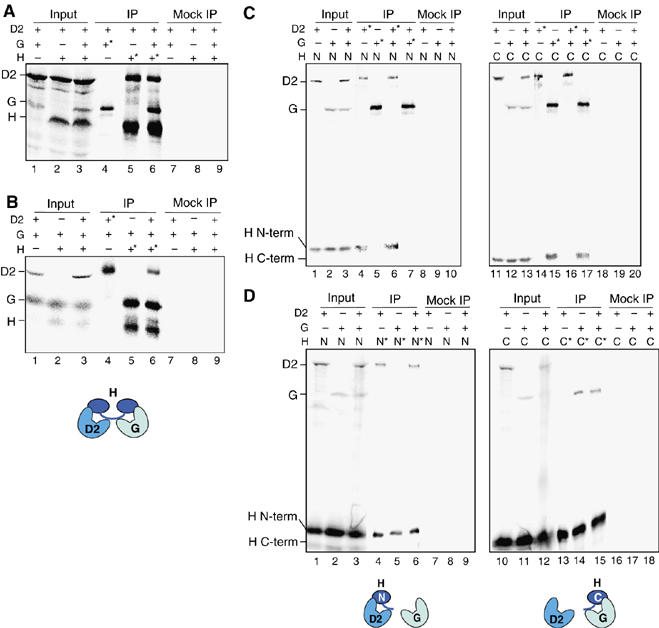
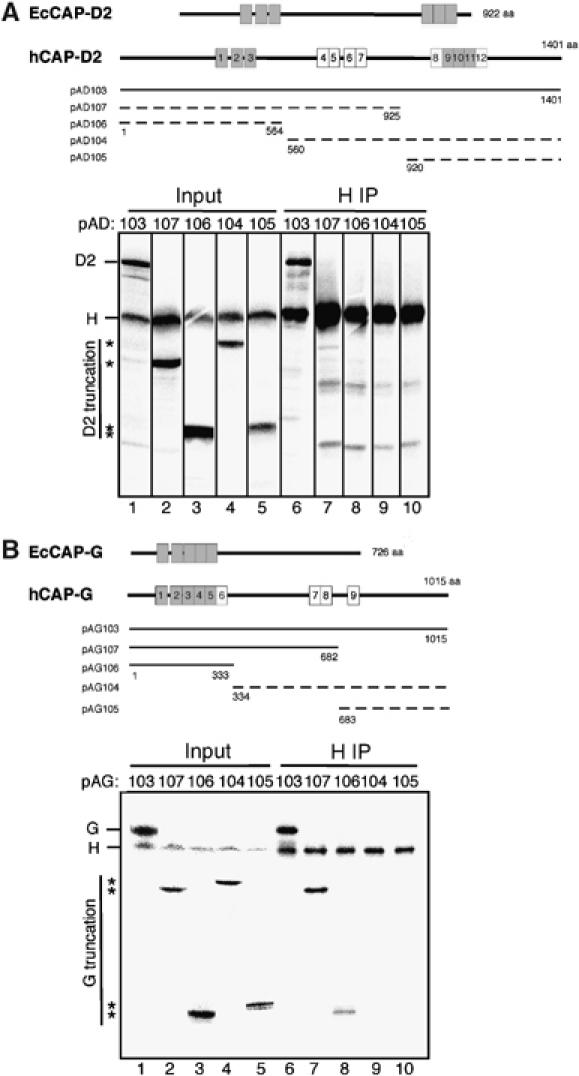
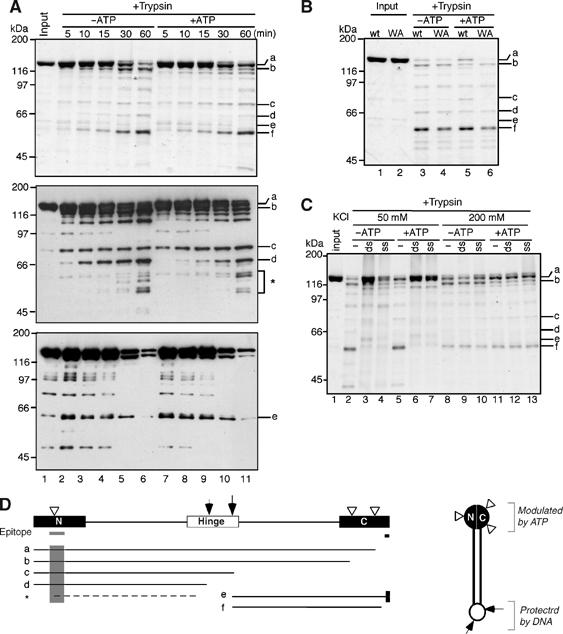
Vertebrate cells possess two different condensin complexes, known as condensin I and condensin II, that play a fundamental role in chromosome assembly and segregation during mitosis. Each complex contains a pair of structural maintenance of chromosomes (SMC) ATPases, a kleisin subunit and two HEAT-repeat subunits. Here we use recombinant human condensin subunits to determine their geometry within each complex. We show that both condensin I and condensin II have a pseudo-symmetrical structure, in which the N-terminal half of kleisin links the first HEAT subunit to SMC2, whereas its C-terminal half links the second HEAT subunit to SMC4. No direct interactions are detectable between the SMC dimer and the HEAT subunits, indicating that the kleisin subunit acts as the linchpin in holocomplex assembly. ATP has little, if any, effects on the assembly and integrity of condensin. Cleavage pattern of SMC2 by limited proteolysis is changed upon its binding to ATP or DNA. Our results shed new light on the architecture and dynamics of this highly elaborate machinery designed for chromosome assembly.
Figures
Similar articles
-
Disturbance in function and expression of condensin affects chromosome compaction in HeLa cells.Cell Biol Int. 2011 Jul;35(7):735-40. doi: 10.1042/CBI20100646. Cell Biol Int. 2011. PMID: 21395557
-
Contribution of hCAP-D2, a non-SMC subunit of condensin I, to chromosome and chromosomal protein dynamics during mitosis.Mol Cell Biol. 2005 Jan;25(2):740-50. doi: 10.1128/MCB.25.2.740-750.2005. Mol Cell Biol. 2005. PMID: 15632074 Free PMC article.
-
Cti1/C1D interacts with condensin SMC hinge and supports the DNA repair function of condensin.Proc Natl Acad Sci U S A. 2004 May 25;101(21):8078-83. doi: 10.1073/pnas.0307976101. Epub 2004 May 17. Proc Natl Acad Sci U S A. 2004. PMID: 15148393 Free PMC article.
-
Condensin and biological role of chromosome condensation.Prog Cell Cycle Res. 2003;5:361-7. Prog Cell Cycle Res. 2003. PMID: 14593730 Review.
-
Condensins: organizing and segregating the genome.Curr Biol. 2005 Apr 12;15(7):R265-75. doi: 10.1016/j.cub.2005.03.037. Curr Biol. 2005. PMID: 15823530 Review.
Cited by
-
Subunits of human condensins are potential therapeutic targets for cancers.Cell Div. 2018 Feb 20;13:2. doi: 10.1186/s13008-018-0035-3. eCollection 2018. Cell Div. 2018. PMID: 29467813 Free PMC article.
-
Condensin, master organizer of the genome.Chromosome Res. 2017 Mar;25(1):61-76. doi: 10.1007/s10577-017-9553-0. Epub 2017 Feb 9. Chromosome Res. 2017. PMID: 28181049 Review.
-
Condensin II mutation causes T-cell lymphoma through tissue-specific genome instability.Genes Dev. 2016 Oct 1;30(19):2173-2186. doi: 10.1101/gad.284562.116. Epub 2016 Oct 13. Genes Dev. 2016. PMID: 27737961 Free PMC article.
-
Structural Basis for a Safety-Belt Mechanism That Anchors Condensin to Chromosomes.Cell. 2017 Oct 19;171(3):588-600.e24. doi: 10.1016/j.cell.2017.09.008. Epub 2017 Oct 5. Cell. 2017. PMID: 28988770 Free PMC article.
-
Human Condensin I and II Drive Extensive ATP-Dependent Compaction of Nucleosome-Bound DNA.Mol Cell. 2020 Jul 2;79(1):99-114.e9. doi: 10.1016/j.molcel.2020.04.026. Epub 2020 May 22. Mol Cell. 2020. PMID: 32445620 Free PMC article.
References
-
- Arumugam P, Gruber S, Tanaka K, Haering CH, Mechtler K, Nasmyth K (2003) ATP hydrolysis is required for cohesin's association with chromosomes. Curr Biol 13: 1941–1953 - PubMed
-
- Cobbe N, Heck MMS (2004) The evolution of SMC proteins: phylogenetic analysis and structural implications. Mol Biol Evol 21: 332–347 - PubMed
-
- Haering CH, Lowe J, Hochwagen A, Nasmyth K (2002) Molecular architecture of SMC proteins and the yeast cohesin complex. Mol Cell 9: 773–788 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
