

Single-channel properties of glycine receptors of juvenile rat spinal motoneurones in vitro
- PMID: 17272347
- PMCID: PMC2075563
- DOI: 10.1113/jphysiol.2006.125740
Single-channel properties of glycine receptors of juvenile rat spinal motoneurones in vitro
Abstract
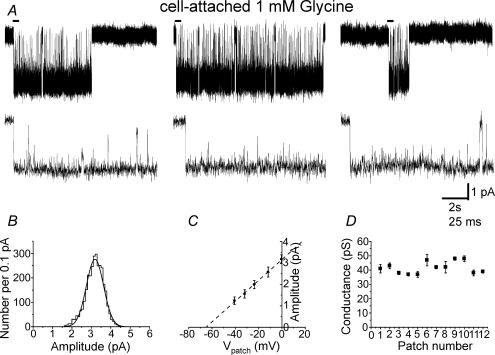
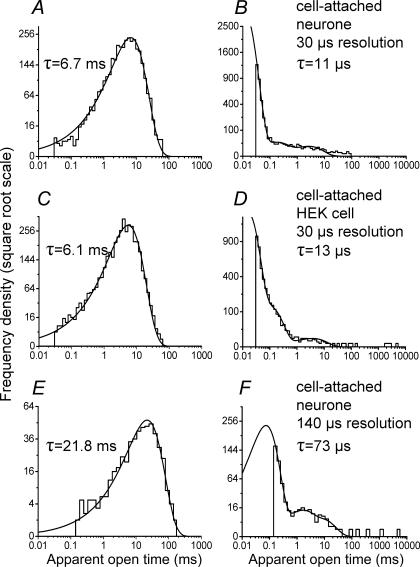
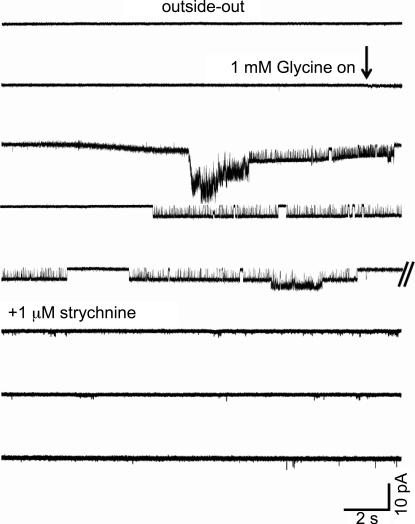
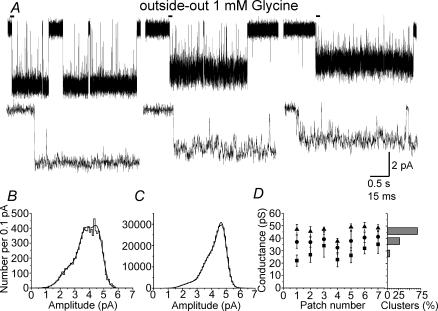
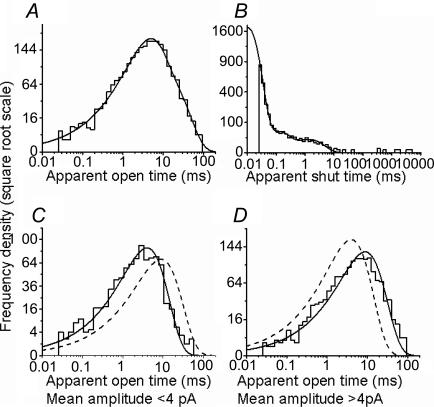
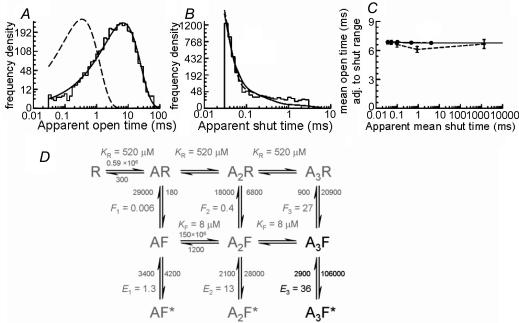
An essential step in understanding fast synaptic transmission is to establish the activation mechanism of synaptic receptors. The purpose of this work was to extend our detailed single-channel kinetic characterization of alpha1beta glycine channels from rat recombinant receptors to native channels from juvenile (postnatal day 12-16) rat spinal cord slices. In cell-attached patches from ventral horn neurones, 1 mM glycine elicited clusters of channel openings to a single conductance level (41 +/- 1 pS, n = 12). This is similar to that of recombinant heteromers. However, fewer than 1 in 100 cell-attached patches from spinal neurones contained glycine channels. Outside-out patches gave a much higher success rate, but glycine channels recorded in this configuration appeared different, in that clusters opened to three conductance levels (28 +/- 2, 38 +/- 1 and 46 +/- 1 pS, n = 7, one level per cluster, all levels being detected in each patch). Furthermore, open period properties were different for the different conductances. As a consequence of this, the only recordings suitable for kinetic analysis were the cell-attached ones. Low channel density precluded recording at glycine concentrations other than 1 mM, but the 1 mm data allowed us to estimate the fully bound gating constants by global model fitting of the 'flip' mechanism of Burzomato and co-workers. Our results suggest that glycine receptors on ventral horn neurones in the juvenile rat are heteromers and have fast gating, similar to that of recombinant alpha1beta receptors.
Figures
References
-
- Beato M, Nistri A. Interaction between disinhibited bursting and fictive locomotor patterns in the rat isolated spinal cord. J Neurophysiol. 1999;82:2029–2038. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials