

Apical GLUT2 and Cav1.3: regulation of rat intestinal glucose and calcium absorption
- PMID: 17272350
- PMCID: PMC2075547
- DOI: 10.1113/jphysiol.2006.124768
Apical GLUT2 and Cav1.3: regulation of rat intestinal glucose and calcium absorption
Abstract
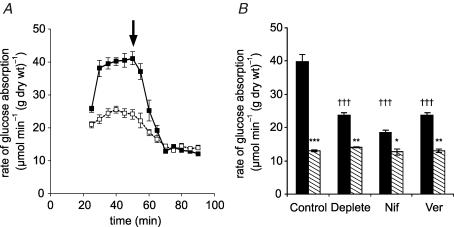
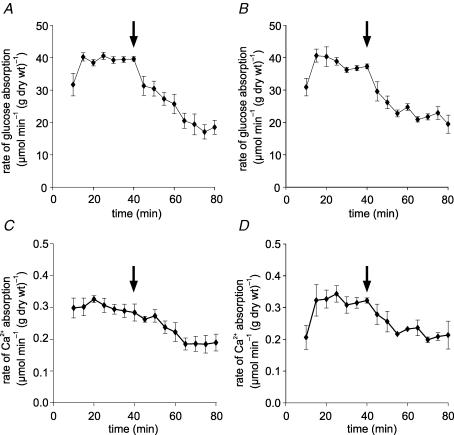
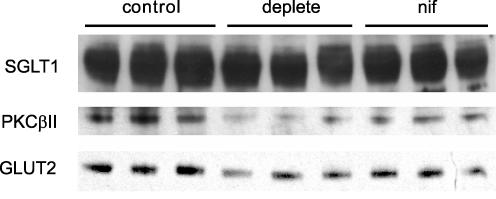
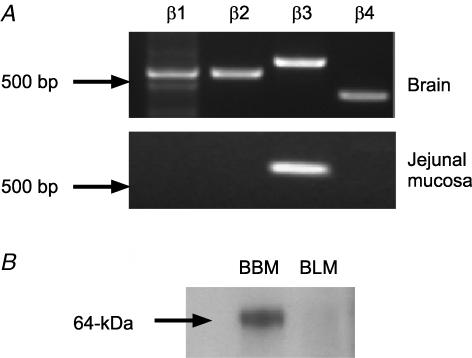
We have proposed a model of intestinal glucose absorption in which transport by SGLT1 induces rapid insertion and activation of GLUT2 in the apical membrane by a PKC betaII-dependent mechanism. Since PKC betaII requires Ca(2+) and glucose is depolarizing, we have investigated whether glucose absorption is regulated by the entry of dietary Ca(2+) through Ca(v)1.3 in the apical membrane. When rat jejunum was perfused with 75 mM glucose, Ca(2+)-deplete conditions, or perfusion with the L-type antagonists nifedipine and verapamil strongly diminished the phloretin-sensitive apical GLUT2, but not the phloretin-insensitive SGLT1 component of glucose absorption. Western blotting showed that in each case there was a significant decrease in apical GLUT2 level, but no change in SGLT1 level. Inhibition of apical GLUT2 absorption coincided with inhibition of unidirectional (45)Ca(2+) entry by nifedipine and verapamil. At 10 mM luminal Ca(2+), (45)Ca(2+) absorption in the presence of 75 mM glucose was 2- to 3-fold that in the presence of 75 mM mannitol. The glucose-induced component was SGLT1-dependent and nifedipine-sensitive. RT-PCR revealed the presence of Ca(v)beta(3) in jejunal mucosa; Western blotting and immunocytochemistry localized Ca(v)beta(3) to the apical membrane, together with Ca(v)1.3. We conclude that in times of dietary sufficiency Ca(v)1.3 may mediate a significant pathway of glucose-stimulated Ca(2+) entry into the body and that luminal supply of Ca(2+) is necessary for GLUT2-mediated glucose absorption. The integration of glucose and Ca(2+) absorption represents a complex nutrient-sensing system, which allows both absorptive pathways to be regulated rapidly and precisely to match dietary intake.
Figures


References
-
- Auchere D, Tardivel S, Gounelle JC, Drueke T, Lacour B. Role of transcellular pathway in ileal Ca2+ absorption: stimulation by low-Ca2+ diet. Am J Physiol. 1998;275:G951–G956. - PubMed
-
- Auchere D, Tardivel S, Gounelle JC, Lacour B. Stimulation of ileal transport of calcium by sorbitol in in situ perfused loop in rats. Gastroenterol Clin Biol. 1997;21:960–966. - PubMed
-
- Baba R, Yamami M, Sakuma Y, Fujita M, Fujimoto S. Relationship between glucose transporter and changes in the absorptive system in small intestinal absorptive cells during the weaning process. Med Mol Morph. 2005;38:47–53. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous