

Inhaled iloprost suppresses the cardinal features of asthma via inhibition of airway dendritic cell function
- PMID: 17273558
- PMCID: PMC1783814
- DOI: 10.1172/JCI28949
Inhaled iloprost suppresses the cardinal features of asthma via inhibition of airway dendritic cell function
Abstract
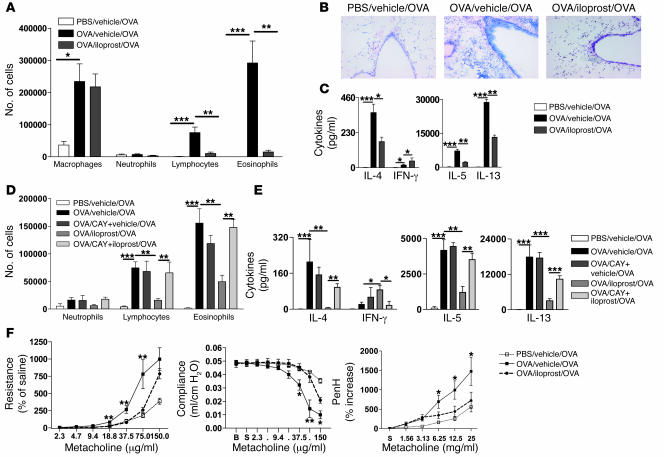
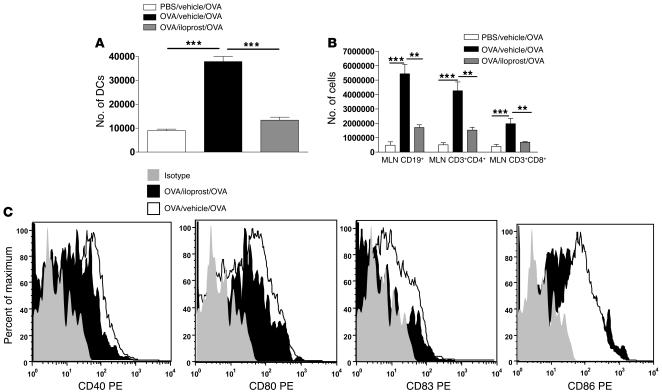
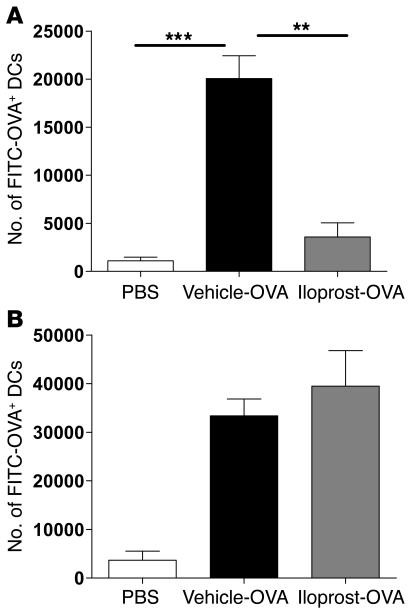
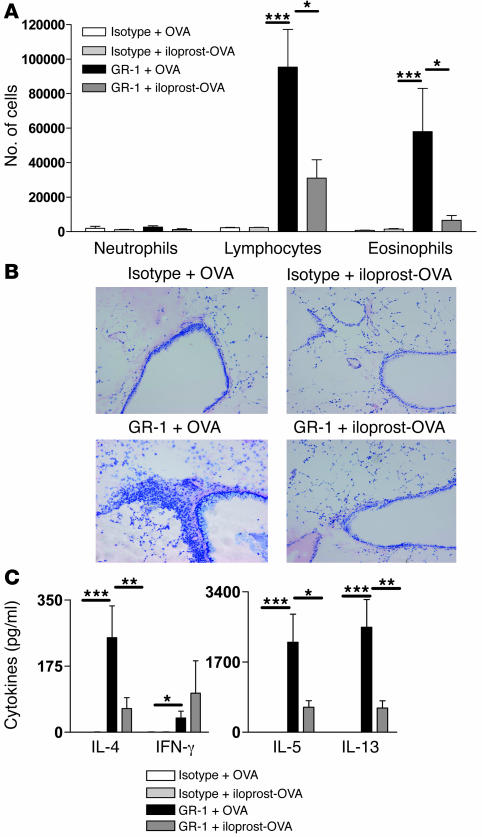
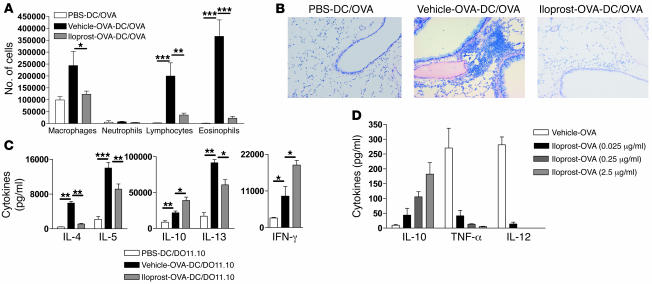
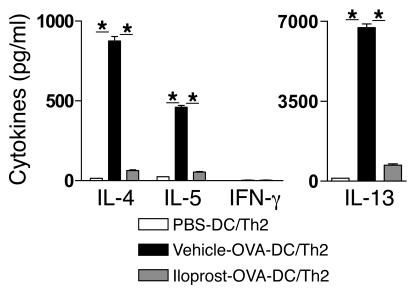
Inhalation of iloprost, a stable prostacyclin (PGI(2)) analog, is a well-accepted and safe treatment for pulmonary arterial hypertension. Although iloprost mainly acts as a vasodilator by binding to the I prostanoid (IP) receptor, recent evidence suggests that signaling via this receptor also has antiinflammatory effects through unclear mechanisms. Here we show in a murine model of asthma that iloprost inhalation suppressed the cardinal features of asthma when given during the priming or challenge phase. As a mechanism of action, iloprost interfered with the function of lung myeloid DCs, critical antigen-presenting cells of the airways. Iloprost treatment inhibited the maturation and migration of lung DCs to the mediastinal LNs, thereby abolishing the induction of an allergen-specific Th2 response in these nodes. The effect of iloprost was DC autonomous, as iloprost-treated DCs no longer induced Th2 differentiation from naive T cells or boosted effector cytokine production in primed Th2 cells. These data should pave the way for a clinical effectiveness study using inhaled iloprost for the treatment of asthma.
Figures
Similar articles
-
Essential role of dendritic cell CD80/CD86 costimulation in the induction, but not reactivation, of TH2 effector responses in a mouse model of asthma.J Allergy Clin Immunol. 2004 Jul;114(1):166-73. doi: 10.1016/j.jaci.2004.03.044. J Allergy Clin Immunol. 2004. PMID: 15241361
-
Local application of FTY720 to the lung abrogates experimental asthma by altering dendritic cell function.J Clin Invest. 2006 Nov;116(11):2935-44. doi: 10.1172/JCI28295. J Clin Invest. 2006. PMID: 17080194 Free PMC article.
-
Essential role of lung plasmacytoid dendritic cells in preventing asthmatic reactions to harmless inhaled antigen.J Exp Med. 2004 Jul 5;200(1):89-98. doi: 10.1084/jem.20040035. J Exp Med. 2004. PMID: 15238608 Free PMC article.
-
Dendritic cells in asthma: a function beyond sensitization.Clin Exp Allergy. 2005 Sep;35(9):1125-34. doi: 10.1111/j.1365-2222.2005.02321.x. Clin Exp Allergy. 2005. PMID: 16164437 Review.
-
Biology of lung dendritic cells at the origin of asthma.Immunity. 2009 Sep 18;31(3):412-24. doi: 10.1016/j.immuni.2009.08.008. Immunity. 2009. PMID: 19766084 Review.
Cited by
-
A prostacyclin analogue, iloprost, protects from bleomycin-induced pulmonary fibrosis in mice.Respir Res. 2010 Mar 20;11(1):34. doi: 10.1186/1465-9921-11-34. Respir Res. 2010. PMID: 20302663 Free PMC article.
-
Prostaglandins in asthma and allergic diseases.Pharmacol Ther. 2019 Jan;193:1-19. doi: 10.1016/j.pharmthera.2018.08.001. Epub 2018 Aug 3. Pharmacol Ther. 2019. PMID: 30081047 Free PMC article. Review.
-
Effects of PGI2 analogues on Th1- and Th2-related chemokines in monocytes via epigenetic regulation.J Mol Med (Berl). 2011 Jan;89(1):29-41. doi: 10.1007/s00109-010-0694-2. Epub 2010 Nov 18. J Mol Med (Berl). 2011. PMID: 21085923
-
Iloprost for asthma.J Mol Med (Berl). 2011 Jan;89(1):1-3. doi: 10.1007/s00109-010-0705-3. J Mol Med (Berl). 2011. PMID: 21128066 No abstract available.
-
Prostaglandin I₂promotes the development of IL-17-producing γδ T cells that associate with the epithelium during allergic lung inflammation.J Immunol. 2011 Nov 15;187(10):5380-91. doi: 10.4049/jimmunol.1101261. Epub 2011 Oct 5. J Immunol. 2011. PMID: 21976777 Free PMC article.
References
-
- Liu M.C., et al. Evidence for elevated levels of histamine, prostaglandin D2, and other bronchoconstricting prostaglandins in the airways of subjects with mild asthma. Am. Rev. Respir. Dis. 1990;142:126–132. - PubMed
-
- Carey M.A., Germolec D.R., Langenbach R., Zeldin D.C. Cyclooxygenase enzymes in allergic inflammation and asthma. Prostaglandins Leukot. Essent. Fatty Acids. 2003;69:157–162. - PubMed
-
- Warner T.D., Mitchell J.A. Cyclooxygenases: new forms, new inhibitors, and lessons from the clinic. FASEB J. 2004;18:790–804. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
