

Intraflagellar transport protein 27 is a small G protein involved in cell-cycle control
- PMID: 17276912
- PMCID: PMC1905864
- DOI: 10.1016/j.cub.2006.12.040
Intraflagellar transport protein 27 is a small G protein involved in cell-cycle control
Abstract
Background: Intraflagellar transport (IFT) is a motility process operating between the ciliary/flagellar (interchangeable terms) membrane and the microtubular axoneme of motile and sensory cilia. Multipolypeptide IFT particles, composed of complexes A and B, carry flagellar precursors to their assembly site at the flagellar tip (anterograde) powered by kinesin, and turnover products from the tip back to the cytoplasm (retrograde) driven by cytoplasmic dynein. IFT is essential for the assembly and maintenance of almost all eukaryotic cilia and flagella, and mutations affecting either the IFT motors or the IFT particle polypeptides result in the inability to assemble normal flagella or in defects in the sensory functions of cilia.
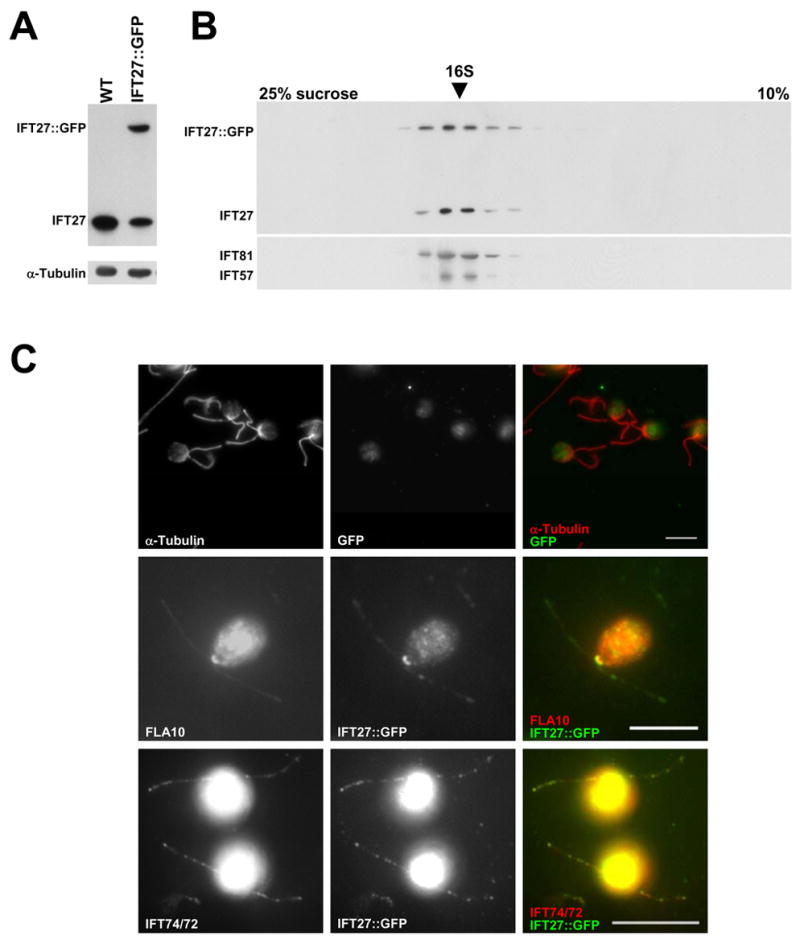
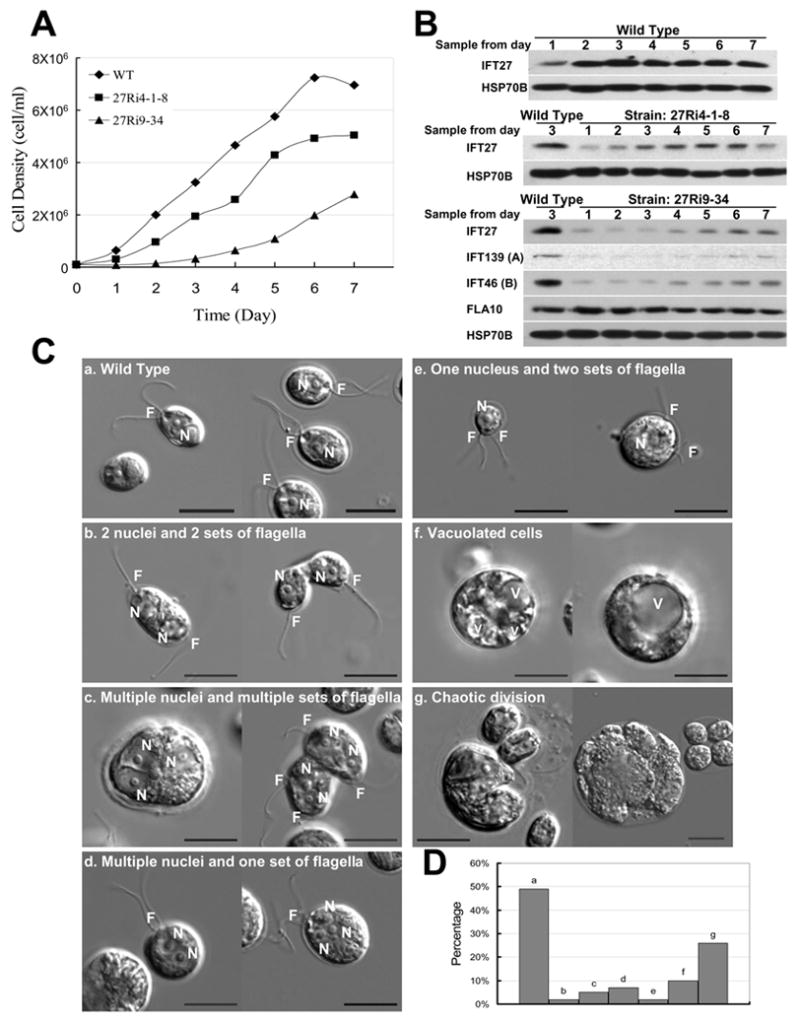
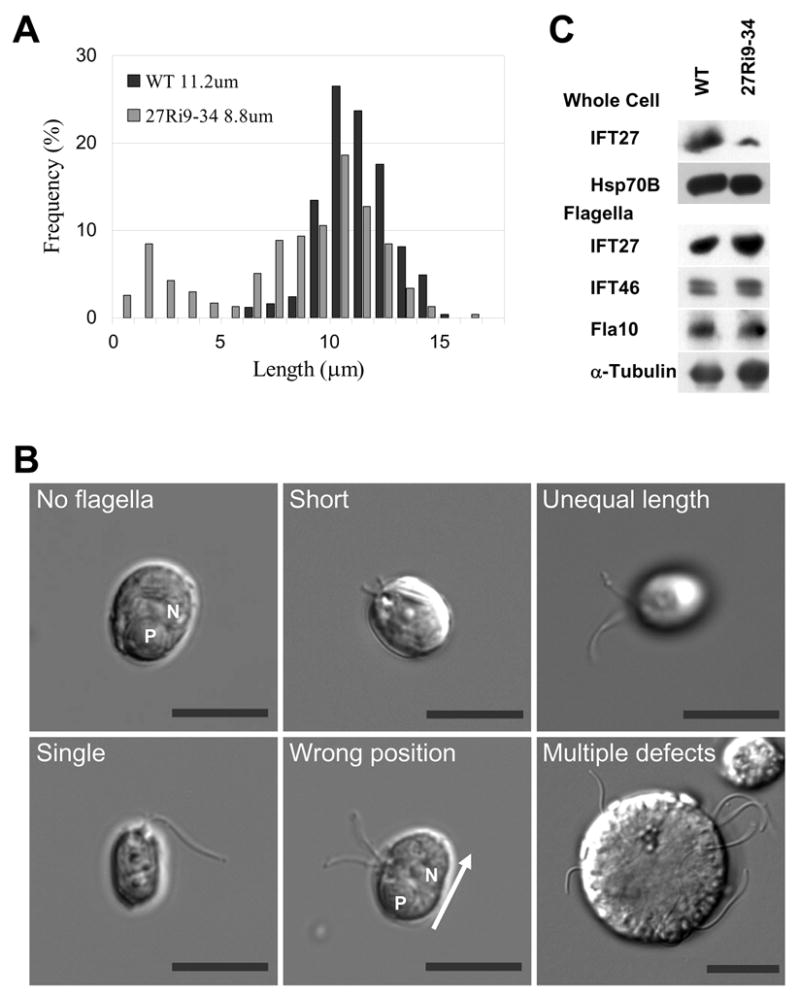
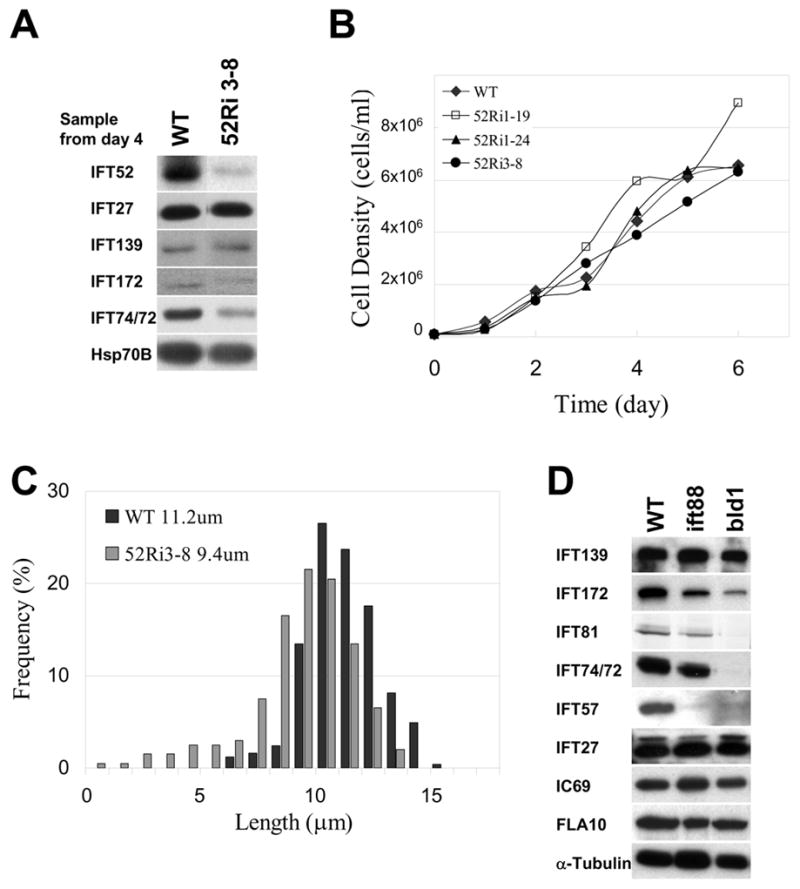
Results: We found that the IFT complex B polypeptide, IFT27, is a Rab-like small G protein. Reduction of the level of IFT27 by RNA interference reduces the levels of other complex A and B proteins, suggesting that this protein is instrumental in maintaining the stability of both IFT complexes. Furthermore, in addition to its role in flagellar assembly, IFT27 is unique among IFT polypeptides in that its partial knockdown results in defects in cytokinesis and elongation of the cell cycle and a more complete knockdown is lethal.
Conclusion: IFT27, a small G protein, is one of a growing number of flagellar proteins that are now known to have a role in cell-cycle control.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
