

Indirect activation of a plant nucleotide binding site-leucine-rich repeat protein by a bacterial protease
- PMID: 17277084
- PMCID: PMC1790868
- DOI: 10.1073/pnas.0608779104
Indirect activation of a plant nucleotide binding site-leucine-rich repeat protein by a bacterial protease
Abstract
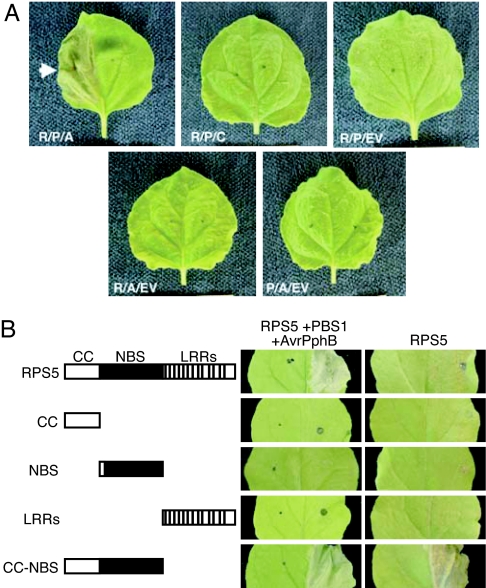
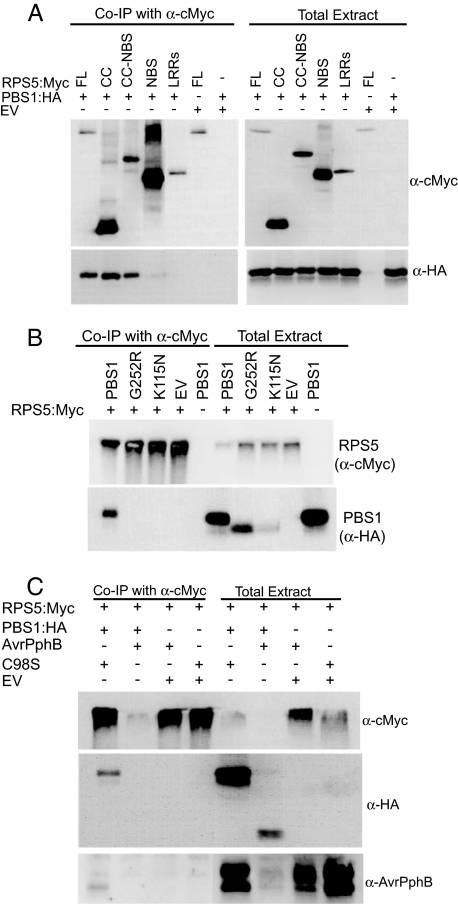
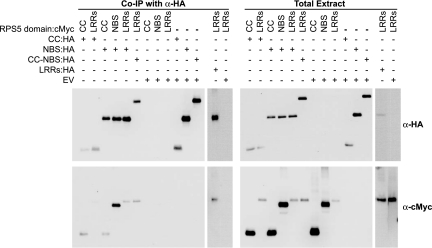
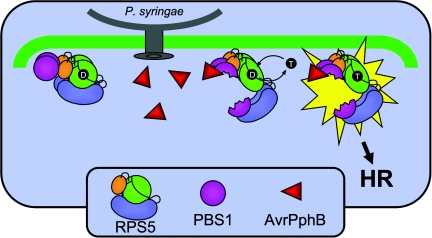
Nucleotide binding site-leucine-rich repeat (NBS-LRR) proteins mediate pathogen recognition in both mammals and plants. The molecular mechanisms by which pathogen molecules activate NBS-LRR proteins are poorly understood. Here we show that RPS5, a NBS-LRR protein from Arabidopsis, is activated by AvrPphB, a bacterial protease, via an indirect mechanism. When transiently expressed in Nicotiana benthamiana leaves, full-length RPS5 protein triggered programmed cell death, but only when coexpressed with AvrPphB and a second Arabidopsis protein, PBS1, which is a specific substrate of AvrPphB. Using coimmunoprecipitation analysis, we found that PBS1 is in a complex with the N-terminal coiled coil (CC) domain of RPS5 before exposure to AvrPphB. Deletion of the RPS5 LRR domain caused RPS5 to constitutively activate programmed cell death, even in the absence of AvrPphB and PBS1, and this activation depended on both the CC and NBS domains. The LRR and CC domains both coimmunoprecipitate with the NBS domain but not with each other. Thus, the LRR domain appears to function in part to inhibit RPS5 signaling, and cleavage of PBS1 by AvrPphB appears to release RPS5 from this inhibition. An amino acid substitution in the NBS site of RPS5 that is known to inhibit ATP binding in other NBS-LRR proteins blocked activation of RPS5, whereas a substitution thought to inhibit ATP hydrolysis constitutively activated RPS5. Combined, these data suggest that ATP versus ADP binding functions as a molecular switch that is flipped by cleavage of PBS1.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Inohara N, Chamaillard M, McDonald C, Nunez G. Annu Rev Biochem. 2005;74:355–383. - PubMed
-
- Jones JD, Dangl JL. Nature. 2006;444:323–329. - PubMed
-
- Shao F, Golstein C, Ade J, Stoutemyer M, Dixon JE, Innes RW. Science. 2003;301:1230–1233. - PubMed
-
- Shao F, Merritt PM, Bao Z, Innes RW, Dixon JE. Cell. 2002;109:575–588. - PubMed
-
- Swiderski MR, Innes RW. Plant J. 2001;26:101–112. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
