

CD4(+)CD25(-)Foxp3(-) Th1 cells are the source of IL-10-mediated immune suppression in chronic cutaneous leishmaniasis
- PMID: 17283207
- PMCID: PMC2118728
- DOI: 10.1084/jem.20061886
CD4(+)CD25(-)Foxp3(-) Th1 cells are the source of IL-10-mediated immune suppression in chronic cutaneous leishmaniasis
Abstract
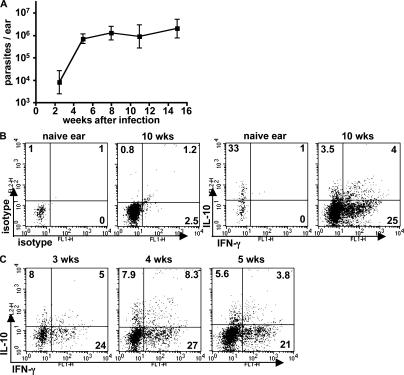
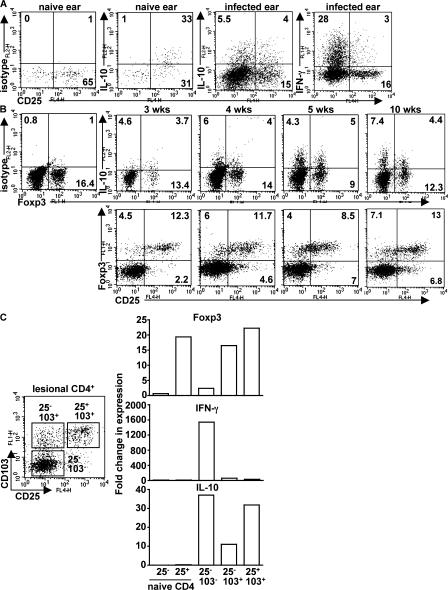
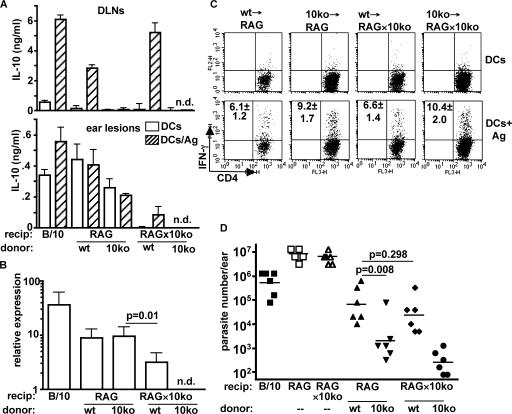
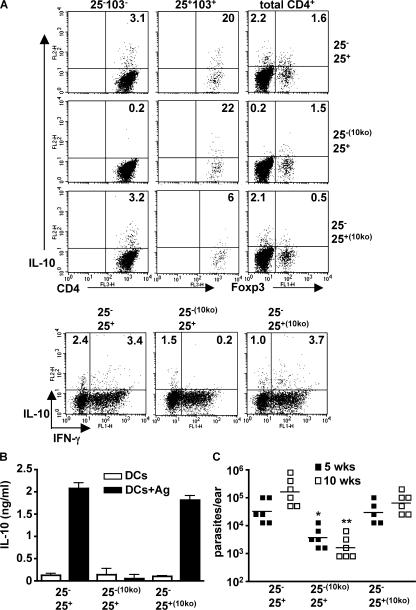
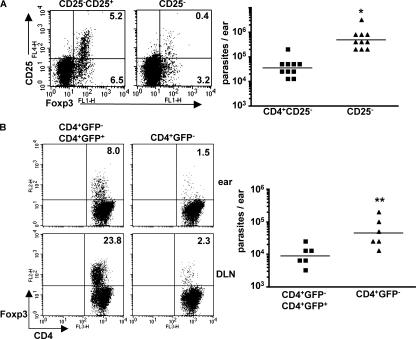
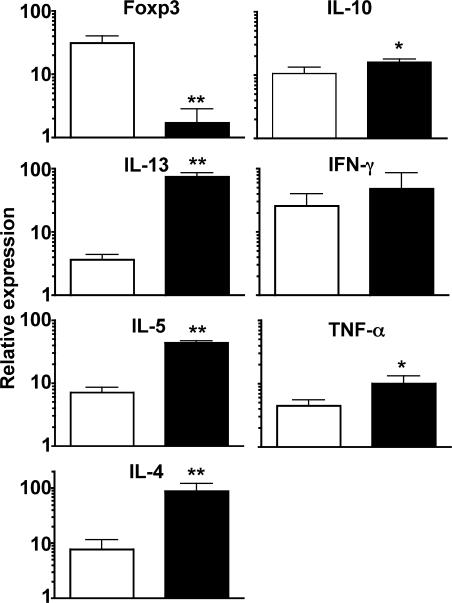
Nonhealing forms of leishmaniasis in humans are commonly associated with elevated levels of the deactivating cytokine IL-10, and in the mouse, normally chronic infections can be cleared in the absence of IL-10. Using a Leishmania major strain that produces nonhealing dermal lesions in a T helper type 1 (Th1) cell-polarized setting, we have analyzed the cellular sources of IL-10 and their relative contribution to immune suppression. IL-10 was produced by innate cells, as well as CD4(+)CD25(+)Foxp3(+) and CD4(+)CD25(-)Foxp3(-) T cells in the chronic lesion. Nonetheless, only IL-10 production by antigen-specific CD4(+)CD25(-)Foxp3(-) T cells, the majority of which also produced IFN-gamma, was necessary for suppression of acquired immunity in Rag(-/-) reconstituted mice. Surprisingly, Rag(-/-) mice reconstituted with naive CD4(+) T cells depleted of natural T regulatory cells developed more severe infections, associated with elevated levels of IL-10 and, especially, Th2 cytokines in the site. The data demonstrate that IL-10-producing Th1 cells, activated early in a strong inflammatory setting as a mechanism of feedback control, are the principal mediators of T cell-derived IL-10-dependent immune suppression in a chronic intracellular infection.
Figures
References
-
- Sacks, D., and N. Noben-Trauth. 2002. The immunology of susceptibility and resistance to Leishmania major in mice. Nat. Rev. Immunol. 2:845–858. - PubMed
-
- Louzir, H., P.C. Melby, A. Ben Salah, H. Marrakchi, K. Aoun, R. Ben Ismail, and K. Dellagi. 1998. Immunologic determinants of disease evolution in localized cutaneous leishmaniasis due to Leishmania major. J. Infect. Dis. 177:1687–1695. - PubMed
-
- Gasim, S., A.M. Elhassan, E.A. Khalil, A. Ismail, A.M. Kadaru, A. Kharazmi, and T.G. Theander. 1998. High levels of plasma IL-10 and expression of IL-10 by keratinocytes during visceral leishmaniasis predict subsequent development of post-kala-azar dermal leishmaniasis. Clin. Exp. Immunol. 111:64–69. (published erratum appears in Clin. Exp. Immunol. 1998. 112:547) - PMC - PubMed
-
- Ismail, A., A.M. El Hassan, K. Kemp, S. Gasim, A.E. Kadaru, T. Moller, A. Kharazmi, and T.G. Theander. 1999. Immunopathology of post kala-azar dermal leishmaniasis (PKDL): T-cell phenotypes and cytokine profile. J. Pathol. 189:615–622. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
