

Restoring species-specific posttransfer editing activity to a synthetase with a defunct editing domain
- PMID: 17283340
- PMCID: PMC1892958
- DOI: 10.1073/pnas.0611110104
Restoring species-specific posttransfer editing activity to a synthetase with a defunct editing domain
Abstract
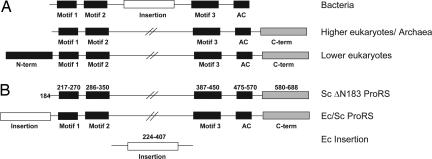
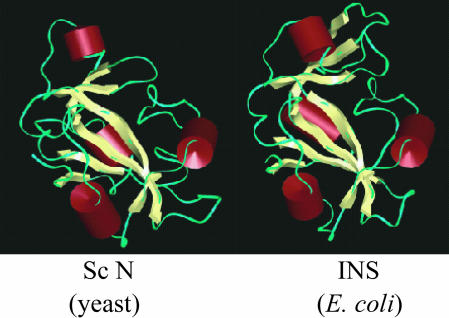
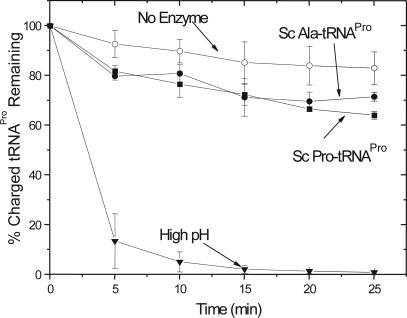
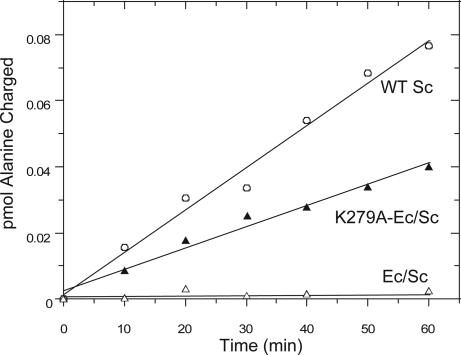
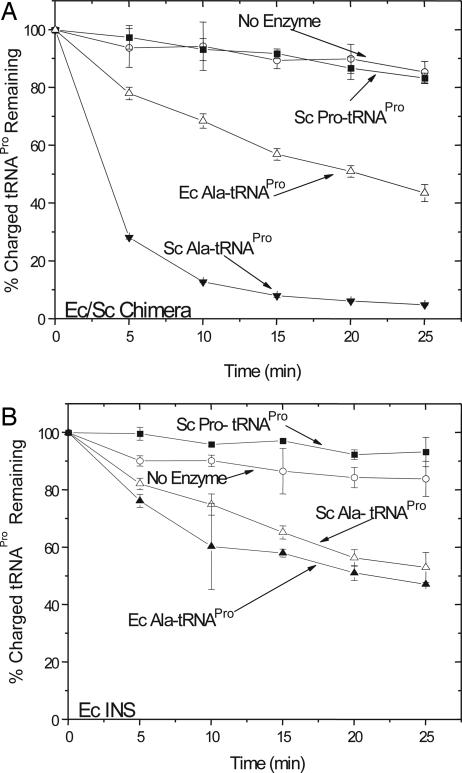
Aminoacyl-tRNA synthetases are multidomain proteins responsible for the attachment of specific amino acids to their tRNA substrates. Prolyl-tRNA synthetases (ProRSs) are notable due to their particularly diverse architectures through evolution. For example, Saccharomyces cerevisiae ProRS possesses an N-terminal extension with weak homology to a bacterial-specific domain typically present as an insertion (INS) within the aminoacylation active site. The INS domain has been shown to contain a "posttransfer" editing active site responsible for cleaving the aminoacyl-ester bond of misacylated Ala-tRNA(Pro) species. However, wild-type S. cerevisiae ProRS does not perform posttransfer editing in vitro. Here, we show that replacement of the N-terminal domain of S. cerevisiae ProRS with the Escherichia coli INS domain confers posttransfer editing function to this chimeric enzyme, with specificity for yeast Ala-tRNA(Pro). In contrast, the isolated INS domain displays only weak editing activity and lacks tRNA sequence specificity. These results emphasize the modular nature of synthetase editing active sites and demonstrate how in evolution, a weak editing activity can be converted to a more robust state through fusion to the body of a synthetase. In this manner, a single editing module can be distributed to different synthetases, and simultaneously acquire specificity and enhanced activity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Berg P. Annu Rev Biochem. 1961;30:293–324.
-
- Fersht AR, Dingwall C. Biochemistry. 1979;18:2627–2631. - PubMed
-
- Lin L, Hale SP, Schimmel P. Nature. 1996;384:33–34. - PubMed
-
- Nureki O, Vassylyev DG, Tateno M, Shimada A, Nakama T, Fukai S, Konno M, Hendrickson TL, Schimmel P, Yokoyama S. Science. 1998;280:578–582. - PubMed
-
- Sankaranarayanan R, Dock-Bregeon AC, Romby P, Caillet J, Springer M, Rees B, Ehresmann C, Ehresmann B, Moras D. Cell. 1999;97:371–381. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
