

Acute postnatal ablation of Hif-2alpha results in anemia
- PMID: 17284606
- PMCID: PMC1892942
- DOI: 10.1073/pnas.0608382104
Acute postnatal ablation of Hif-2alpha results in anemia
Abstract
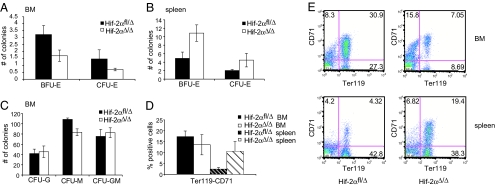
Adaptive transcriptional responses to oxygen deprivation (hypoxia) are mediated by the hypoxia-inducible factors (HIFs), heterodimeric transcription factors composed of two basic helix-loop-helix-PAS family proteins. The transcriptional activity of HIF is determined by the hypoxic stabilization of the HIF-alpha proteins. HIF-1alpha and HIF-2alpha exhibit high sequence homology but have different mRNA expression patterns; HIF-1alpha is expressed ubiquitously whereas HIF-2alpha expression is more restricted to certain tissues, e.g., the endothelium, lung, brain, and neural crest derivatives. Germ-line deletion of either HIF subunit is embryonic lethal with unique features suggesting important roles for both HIF-alpha isoforms. Global deletion of Hif-2alpha results in distinct phenotypes depending on the mouse strain used for the mutation, clearly demonstrating an important role for HIF-2alpha in mouse development. The function of HIF-2alpha in adult life, however, remains incompletely understood. In this study, we describe the generation of a conditional murine Hif-2alpha allele and the effect of its acute postnatal ablation. Under very stringent conditions, we ablate Hif-2alpha after birth and compare the effect of acute global deletion of Hif-2alpha and Hif-1alpha. Our results demonstrate that HIF-2alpha plays a critical role in adult erythropoiesis, with acute deletion leading to anemia. Furthermore, although HIF-1alpha was first purified and cloned based on its affinity for the human erythropoietin (EPO) 3' enhancer hypoxia response element (HRE) and regulates Epo expression during mouse embryogenesis, HIF-2alpha is the critical alpha isoform regulating Epo under physiologic and stress conditions in adults.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Schofield CJ, Ratcliffe PJ. Nat Rev Mol Cell Biol. 2004;5:343–354. - PubMed
-
- Semenza GL. Trends Mol Med. 2001;7:345–350. - PubMed
-
- Ivan M, Kondo K, Yang H, Kim W, Valiando J, Ohh M, Salic A, Asara JM, Lane WS, Kaelin WG., Jr Science. 2001;292:464–468. - PubMed
-
- Jaakkola P, Mole DR, Tian YM, Wilson MI, Gielbert J, Gaskell SJ, Kriegsheim A, Hebestreit HF, Mukherji M, et al. Science. 2001;292:468–472. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
